

Understanding the Invasion, Ecological Adaptations, and Management Strategies of Bactrocera dorsalis in China: A Review


1
Department of Entomology, College of Plant Protection, South China Agricultural University, Guangzhou 510642, China
2
Xinjiang Institute of Ecology and Geography, Urumqi 830011, China
*
Author to whom correspondence should be addressed.
Horticulturae 2023, 9(9), 1004; https://doi.org/10.3390/horticulturae9091004
Submission received: 18 July 2023
/
Revised: 25 August 2023
/
Accepted: 1 September 2023
/
Published: 5 September 2023
(This article belongs to the Special Issue Orchard Management under Climate Change)
Abstract
:Bactrocera dorsalis (Hendel, 1912) (Diptera: Tephritidae), commonly known as the oriental fruit fly, is a highly destructive pest that globally infests fruits and vegetables, resulting in significant annual economic losses. Initially detected in Taiwan Island, it has rapidly expanded its distribution range to various regions in mainland China since the 1980s, with a continuous northward spread. To mitigate the damage caused by this pest, extensive efforts have been undertaken to comprehend its ecological and physiological adaptations and develop management strategies. This review article provides an overview of the invasion history of B. dorsalis in China, its ecological and physiological mechanisms facilitating its invasion, and the progress made in understanding its major biological characteristics. Moreover, the key approaches for managing B. dorsalis that have been or are likely to be implemented in China are presented, including quarantine measures, monitoring procedures, physical controls, biological controls, the sterile insect technique, RNA interference, and CRISPR-Cas-9. Finally, some suggestions for future research directions are provided.
1. Introduction
Tephritid fruit flies are an economically significant pest species globally, including mainland China [1]. They exhibit endophagous feeding behavior, which causes both quantitative and qualitative yield reductions. As a result, they pose significant threats to global fruit and vegetable production [2,3]. The pest affects a broad array of fruit and fleshy vegetable crops in tropical and subtropical regions. The presence of these pests was first observed in Taiwan Island, China, in 1912 [4,5]. The genus Bactrocera, which comprises a minimum of 440 species [6], is primarily distributed throughout tropical Asia, Australia, and the South Pacific [7,8]. The wide host range, great climate tolerance, and strong dispersing capacities of these species have led to their spread over the Asia Pacific region in the last century, covering all of South-East Asia from India to Hawaii [7]. The oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae), is recognized as a destructive and persistent fruit fly pest. B. dorsalis has been documented to infest over 250 host plant species [9,10], including mango (Mangifera indica L., Anacardiaceae), banana (Musa spp., Musaceae), guava (Psidium guajava L., Myrtaceae), orange (Citrus spp., Rutaceae), papaya (Carica papaya L., Caricaceae), peach (Prunus persica (L.) Batsch, Rosaceae), grape (Vitis spp., Vitaceae), pomegranate (Punica granatum L., Lythraceae), lychee (Litchi chinensis Sonn., Sapindaceae), and longan (Dimocarpus longan Lour., Sapindaceae) [11,12]. Numerous studies have documented the economic damage caused by B. dorsalis. For instance, a study carried out in Thailand found that B. dorsalis infestation in mango farms caused an average annual yield loss of 15.5% [13]. Similarly, in India, fruit fly infestation led to a reduction in the marketable yield of mango by 25–30% [14]. According to an estimate, guava, sapota, citrus fruits, and mango in India, incurred losses equivalent to USD 356 million [15]. This significant economic loss is attributed to the fruit damage caused by B. dorsalis, which can affect 30% to 100% of fruits, depending on the season [16].
In addition to yield reduction, B. dorsalis also leads to the quality degradation of fruits, causing phytosanitary issues and triggering trade restrictions, thereby aggravating economic losses. A study conducted in Taiwan revealed that the infestation of fruit flies resulted in trade restrictions on the export of guava to the United States and Japan, leading to an estimated economic loss of USD 2.5 million per year [17]. These studies demonstrate the substantial economic losses caused by B. dorsalis and emphasize the necessity for implementing effective management strategies to mitigate the impact of this insect pest on horticultural crops. In China, the economic losses caused by the fruit fly pest species in citrus orchards have been widely reported, especially in Guangdong [18] and Fujian Provinces of China [1]. B. dorsalis exhibits three to eleven generations per year in China, with the majority of areas experiencing four to eight generations [8,19]. In the near future, there is the potential for B. dorsalis to expand into temperate northern and southern areas of China [20] (Figure 1).
Keeping in view the severity of the pest, the key objective of the current study is to review the invasion of B. dorsalis from Taiwan to mainland China and its subsequent expansion. The review will also analyze the biological traits of B. dorsalis that have facilitated the insect pest’s invasion into new areas. Moreover, the review will provide an overview of the integrated pest management (IPM) strategies employed in China to control B. dorsalis, encompassing quarantine measures, surveillance techniques, physical and biological control methods, the sterile insect technique, RNA interference, and CRISPR-Cas-9.
2. Dispersion Ecology
2.1. Dispersion of B. dorsalis in China
B. dorsalis was first recorded in Taiwan Island and subsequently invaded Hainan Island, China, in 1934 [21]. Prior to the 1970s, the species was only rarely detected in isolated areas in southern China. However, in the 1980s, its population size increased, due to international trade, resistance to pesticides, abundant host plants, and a lack of natural enemies (predators and parasitoids), and its distribution area expanded to encompass most of southern China. In recent years, B. dorsalis has continued to spread, crossing the Yangtze River and other neighboring areas [22,23]. To predict the potential geographical distribution of B. dorsalis in China, several techniques have been employed. An analysis based on the CLIMEX model has indicated that the species’ most suitable habitats are located in Southern China, including Guangdong, Hainan, and Guangxi Provinces, as well as in Eastern China, including Fujian, Zhejiang, and Shanghai [19,24]. The analysis revealed that moderately suitable habitats for the oriental fruit fly were identified in Southwest China, including Yunnan, Sichuan, and Guizhou Provinces. In contrast, Hunan, Hubei, and Jiangxi Provinces were found to exhibit relatively low suitability. Furthermore, areas located to the north of the Yangtze River were deemed unsuitable for the species [7,8]. Subsequent analyses using the genetic algorithm for rule-set prediction (GARP) ecological niche modeling [25] and the emergence rate model combined with ArcGIS yielded similar predictions [7,24]. So far, B. dorsalis has effectively colonized the majority of the aforementioned regions and provinces, encompassing territories beyond the Yangtze River and including Henan and Anhui Provinces. Anticipated future developments suggest a substantial northward expansion of its range [26] (Figure 2). The utilization of microsatellite loci analysis unveiled valuable insights into the origin of oriental fruit fly populations within China, Korea, Thailand, and Laos [27,28]. These regions are believed to have been among the initial areas to be invaded by the species. Furthermore, indications suggest that Guangxi Province could potentially represent another early region of B. dorsalis invasion. The observed genetic differentiation in the hierarchical model using nad1, nad5, Cytb, and concatenated sequences among the various populations of B. dorsalis in China exhibited substantial levels, implying that this invasive pest might have entered China through two distinct invasion routes. The initial route likely originated in Southeast China, leading to subsequent spread into the provinces of Fujian, Taiwan, Guangdong, and Hainan. The second route is thought to have originated in Southwest China, resulting in subsequent dissemination into the provinces of Sichuan, Yunnan, Guangxi, Guizhou, and Hunan, as corroborated by the findings presented in previous studies [29,30].
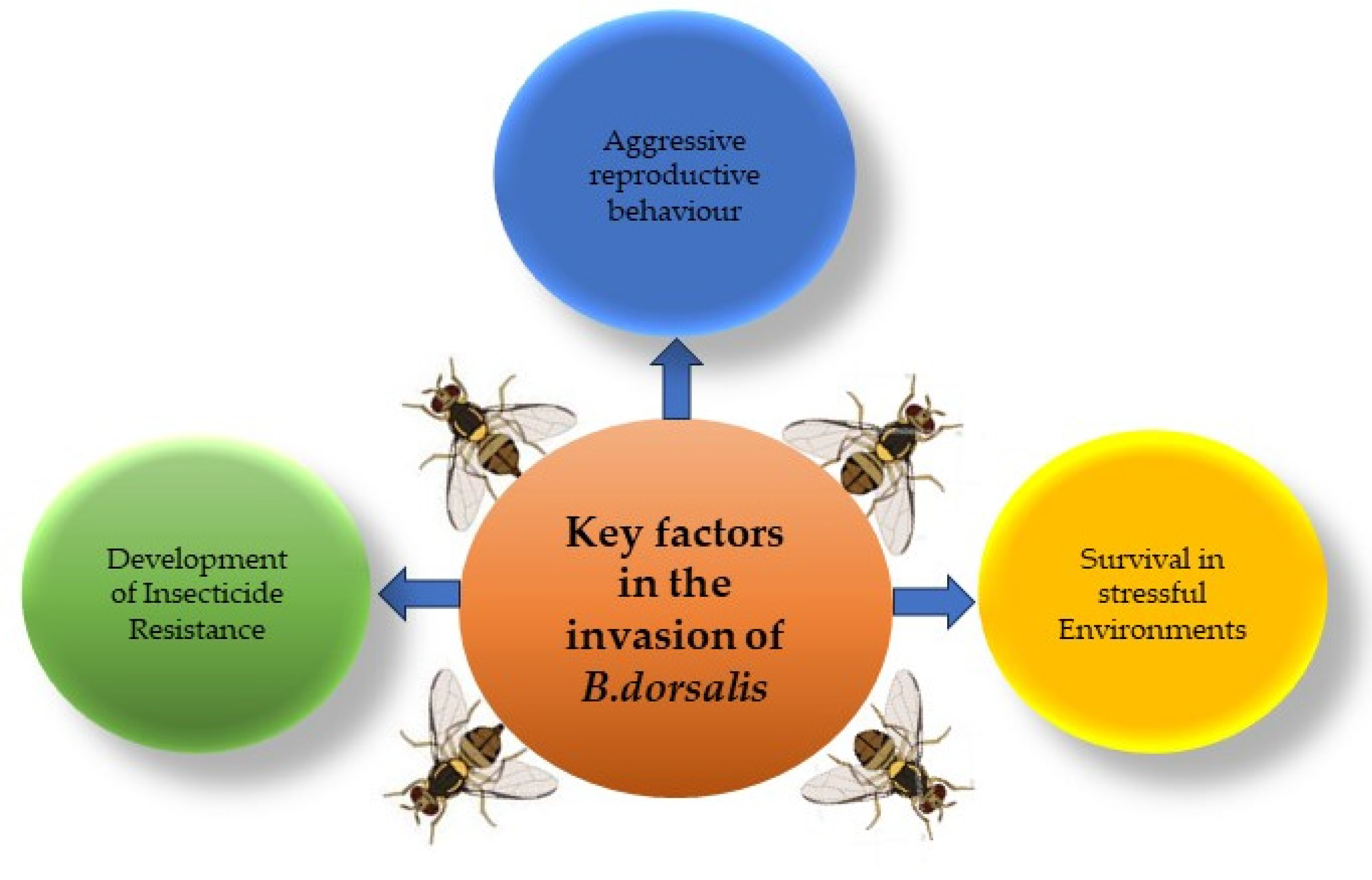
2.2. Key Factors in the Invasion of B. dorsalis
Over the past few decades, numerous research studies have been published on B. dorsalis, with a particular focus on understanding key contributors that help the pest to invade different regions. These studies have investigated various aspects of the insect, including its aggressive reproductive behavior, tolerance to stressful ecological niches, and resilience to chemical pesticides. By examining these factors, researchers aim to understand the mechanisms driving the successful invasion of B. dorsalis and develop effective management strategies. Following is a detailed review of all the factors that make B. dorsalis a highly invasive insect, particularly in China.
2.2.1. Aggressive Reproductive Behavior
One of the reasons for the successful invasion of B. dorsalis is attributed to its reproductive behavior. The pest shows a unique phenomenon of remating in female flies [31], which enables the pest to acquire supplementary material from male accessory gland products E-coniferyl alcohol (ECF) and dimethoxyphenol (DMP), particularly when males have fed on methyl eugenol (ME). These sequestered components can enhance the overall fitness of female flies and contribute to their successful establishment in new environments. The phenomena have also been observed in female B. dorsalis [32,33]. Studies have found that female B. dorsalis, due to multiple mating, tend to have increased fecundity. One study reported that approximately 50% of the B. dorsalis population re-mated and exhibited significantly higher fecundity compared to single-mated females [34]. The remating potential could play a significant role in the rapid population growth of B. dorsalis in natural habitats [35,36]. Group housing, which refers to the social living and interaction of insects, is another reproductive behavior that also plays a role in reproductive success. Group housing has been shown to enhance mating frequency and success in arthropods [37]. Recently, studies have shown that group housing has a positive impact on mating and chemical cue sensitization in several insect species, including B. dorsalis [38,39]. Studies have reported that group-housed flies exhibit higher mating rates compared to those housed in individual containers. Additionally, group housing creates a more natural social environment for the flies, which stimulates reproductive behavior and increases the chances of successful mating. Furthermore, this social interplay has been found to increase the sensitivity of B. dorsalis to chemical cues [40]. This is important for the development of new management options, as chemical cues are often used to attract and trap fruit flies in the field. In group-housed insects, exposure to the pheromones and other volatile chemicals released by conspecifics leads to an increased sensitivity to these cues, which could enhance their efficacy in trapping and monitoring programs. Additional research is needed to fully understand the underlying mechanisms of social interactions, as well as their practical implications for fruit fly management [39]. In another study, the effect of cis-vaccenyl acetate (cVA), a pheromone commonly found in Drosophila [41], on the mating behavior of B. dorsalis was investigated. These results showed that low levels of cVA had a positive impact on the mating rate of B. dorsalis [39], whereas high levels had an inhibitory effect. This finding implies that cVA may play a role in regulating mating behavior not only in Drosophila but also in other insect species [42].
Another study has found that group housing conditions had a positive impact on the mating rates and cVA-mediated behaviors of fruit flies [43]. These effects were shown to be dependent on the activity of the CREB (cAMP response element-binding protein) binding protein (CBP), which regulates gene expression and interacts with CREB. In other words, the presence of other insects in the group setting influences the responses of B. dorsalis to cVA, leading to altered mating rates and behaviors [44]. This study highlights the potential benefits of group housing for enhancing mating behaviors and cVA-mediated responses in B. dorsalis [45]. Further research may be needed to fully understand the mechanisms behind these effects and their practical implications in fruit fly management programs (Figure 3).
2.2.2. Survival in Stressful Environments
B. dorsalis is a highly adaptable species, capable of thriving in a broad geographical range and a diverse set of environmental conditions [46]. With reference to the temperature fluctuations, B. dorsalis demonstrates a remarkable tolerance, with its eggs displaying a high level of resistance to high temperatures and its pupae exhibiting a remarkable ability to withstand low temperatures, especially at the pre-overwintering stage [8,47]. The cold hardiness of future generations of B. dorsalis can be influenced by the host plant they feed on during their larval stage [48]. Physiological mechanisms responsible for the tolerance of B. dorsalis to extreme temperatures have been the subject of investigation. Studies suggest that a multitude of oxidoreductases, binding proteins, and transferases are present in large amounts in adults subjected to high and/or low temperatures, providing a form of physiological protection [49,50]. Moreover, antioxidant enzymes, superoxide dismutase (SODs), likely play a key role in mitigating oxidative damage in B. dorsalis under thermally stressed conditions [51,52]. Also, B. dorsalis can survive in a wide range of humidity levels, with its larvae and pupae thriving and developing in a broad range of moisture content [53]. The larvae of B. dorsalis exhibit a remarkable ability to reduce their weight within two hours when exposed to a dry environment, which highlights its resilience under severe conditions. This rapid response to changes in their environment sets them apart from other insect species. On the other hand, the pupae of B. dorsalis show an ability to survive in wet environments. In a study, it was found that more than 50% of the B. dorsalis pupated in soils with moisture levels between 80 and 100% [54]. Despite the low adult emergence from the highly moist soils, the pupation in highly wet environments shows the resilience of the developmental stages to highly stressful conditions [54]. Thus, the survival of B. dorsalis under stressful regimes serves as one of the important reasons for its success.
2.2.3. Development of Insecticide Resistance
The management of B. dorsalis is typically achieved through chemical insecticides. However, the extended and frequent use of certain synthetic chemicals has led to the evolution of high levels of insecticide resistance in this species, resulting in more destructive outbreaks [55,56]. For example, studies have shown that populations of the oriental fruit fly in Guangdong, China, as well as in many other parts of the world, developed a high level of resistance to trichlorfon between 2007 and 2020 [8,56,57]. In recent years, resistance to other insecticides such as malathion, β-cypermethrin, and abamectin has also been observed [58,59]. The situation has become particularly critical in Hubei, China, where high resistance to cyantraniliprole, a newly developed anthranilic diamide insecticide, has been reported for many insects [60,61]. Various studies have been conducted to investigate the underlying mechanisms and biochemical processes involved in insecticide resistance development. Experiments have revealed that resistance in B. dorsalis is primarily due to increased detoxification mechanisms, including upregulation of cytochrome P450 monooxygenases (P450s), glutathione S-transferases (GSTs), carboxylesterase (CarEs), and insensitivity of acetylcholinesterase (AChE) [62,63]. In addition, physiological resistance, characterized by elevated digestive enzyme activities, has been implicated in resistance development in B. dorsalis [55]. Furthermore, the structural features of the cuticle and the interspace between epidermal cells might play a role in the cuticular penetration of insecticides [58]. Advances in molecular techniques, including heterologous expression and RNA interference, have facilitated the functional characterization of various resistance-related genes in B. dorsalis. For instance, overexpression of three esterase genes has been linked to malathion resistance in this insect species [64,65]. The rapid development of insecticide resistance in B. dorsalis represents one of the important reasons for its success, posing a major challenge to current pest management strategies and necessitating the implementation of innovative approaches for its effective control.
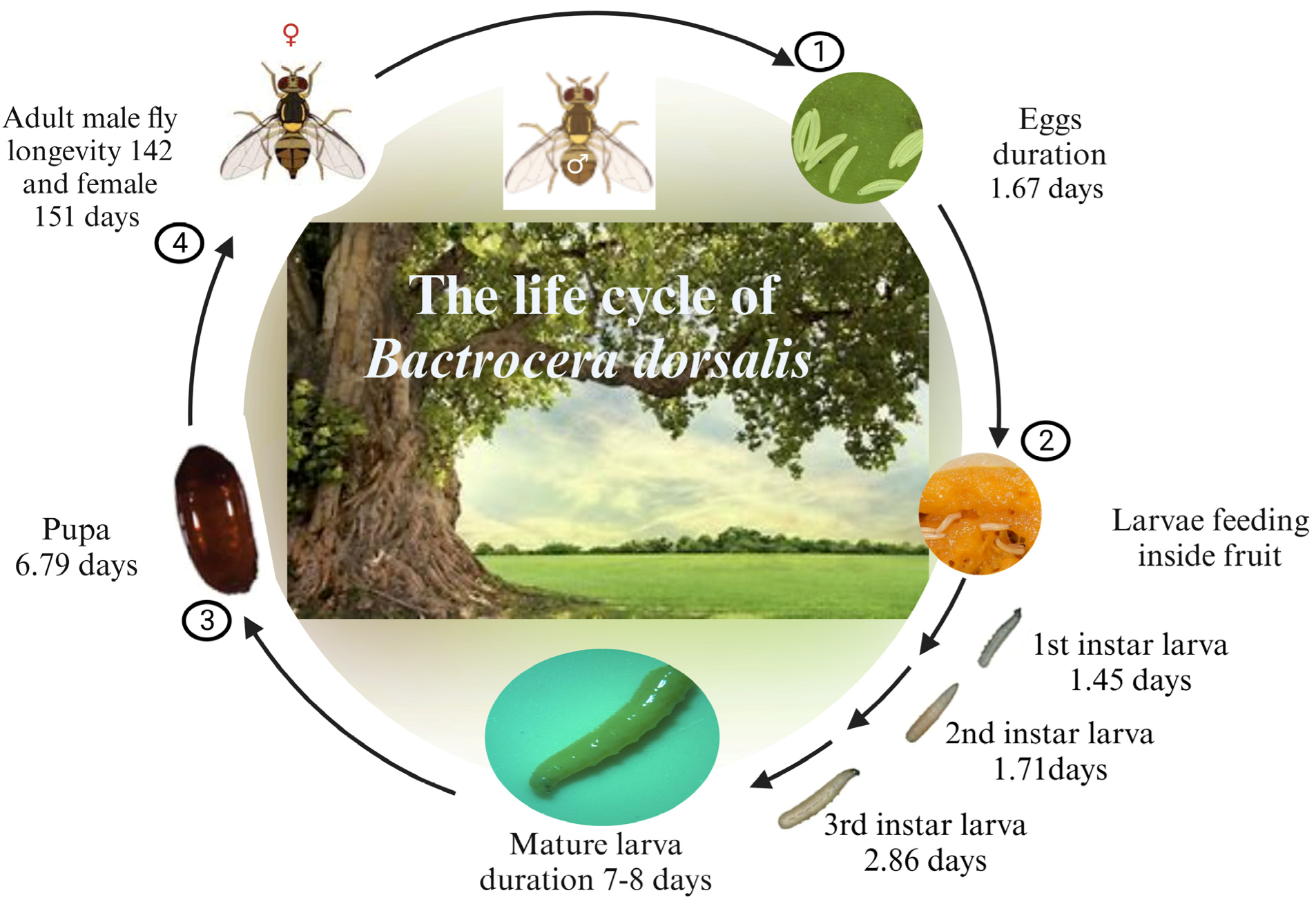
3. Research on B. dorsalis Biology in China
Development studies of B. dorsalis have focused on various life stages, characterizing the developmental period, growth rate, and morphological changes [66,67]. The reproductive biology of this insect has also been extensively studied, focusing on mating behavior, female remating, egg-laying behavior, and egg survival [68,69]. Regarding chemical resistance, the evolution of high levels of insecticide resistance in B. dorsalis has been documented in recent years [8,70]. The biochemical and molecular mechanisms underlying this resistance have been extensively studied. Research has focused on the increased detoxification by cytochrome P450 monooxygenases (P450s), glutathione S-transferases (GSTs), and carboxylesterase (CarEs) [59,71,72]. Physiological resistance, specifically high digestive enzyme activities, has also been implicated in resistance development. Furthermore, recent research has characterized resistance-related genes using heterologous expression and RNA interference techniques [62]. Studies on the associations between B. dorsalis and symbionts have also been conducted to determine their effects on the insect’s fitness and survival. For instance, research has revealed the benefits of gut symbionts in nutrient acquisition and defense against pathogens [73], expanding our understanding of the complex interactions between B. dorsalis and its symbionts. Research on B. dorsalis in China has provided valuable insights into the development, reproduction, chemical resistance, communication, and symbiotic associations of this economically important pest.
3.1. Exploration of Molecular Mechanisms
To gain insights into the molecular mechanisms underlying this regulation, researchers have cloned several key genes involved in the ecdysone biosynthesis pathway from this insect. Specifically, the cloning of ecdysteroid biosynthetic genes Cyp302a1, Cyp315a1, and Cyp314a1, along with the ecdysone receptor gene EcR-B1, has significantly advanced our knowledge in this area [74,75]. The cloning of these genes has facilitated a deeper understanding of how ecdysteroids, specifically 20-hydroxyecdysone (20E), regulate the development of fruit flies. The 20E hormone interacts with the EcR-B1 receptor, which activates a series of downstream target gene expression cascades, leading to cellular processes such as cell proliferation, differentiation, and apoptosis. The genes involved in this process serve as potential molecular targets for the control of this pest [76,77]. The identification of these genes highlights the importance of ecdysteroid signaling in oriental fruit fly development and opens possibilities for utilizing these targets in pest management.
The cloning and characterization of chitin metabolic pathway genes have received significant attention in recent years in B. dorsalis research. Key enzymes in the chitin biosynthesis pathway, namely chitin synthases CHS1 and CHS2, have been cloned and their expression profiles analyzed [78]. Among them, a specific splicing variant of CHS1, CHS1a, shows prominent expression during the larval–pupal and pupal–adult transitions. Knockdown of CHS1a expression using RNA interference (RNAi) results in significant phenotypic defects and mortality in treated larvae [78]. Furthermore, the CHS2 transcript is predominantly found in the larval midgut [79], and its expression is positively correlated with the total chitin content during the insect’s development. Other crucial enzymes involved in the chitin biosynthesis pathway, such as glucose-6-phosphate isomerase and UDP-N-acetylglucosamine pyrophosphorylase, have also been cloned and characterized in drosophila [80]. RNAi-mediated knockdown of their expression leads to larval death and abnormal phenotypes. Moreover, chitinase, an enzyme responsible for chitin degradation and starvation, has been identified and characterized in B. dorsalis as well [81]. Moreover, the significance of the bHLH transcription factor gene, pipsqueak, in the proper regulation of molting in fruit flies has been demonstrated [82]. In the context of cell growth and metabolism regulation, B. dorsalis has an identified ortholog of the target of rapamycin (TOR), a crucial kinase [83]. Furthermore, the evaluation of heat shock proteins 83 and 90 (Hsp 83, 90) has revealed its role in stress response and stress resistance, fecundity, and longevity as its gene expression is upregulated following exposure [84,85]. Furthermore, studies have interpreted the regulation of ecdysteroid hormone biosynthesis in B. dorsalis. The ecdysone biosynthetic gene, cytochrome P450 monooxygenase (Cyp307a1), was identified, and its expression has been shown to be regulated by 20-hydroxyecdysone (20E) and juvenile hormone (JH) [86]. Moreover, the presence of the hormone ecdysone and its receptor, ecdysone receptor, has been confirmed in B. dorsalis [87,88].
In addition to these findings, research has demonstrated the crucial role of digestive enzymes in the growth and development of B. dorsalis. Fourteen genes encoding digestive enzymes, including alpha-amylases, proteases, and chitinases, have been successfully cloned [89]. It has been observed that these enzymes are regulated by 20E, JH, and insulin signaling pathways, further emphasizing their significance in the development of the mosquito and oriental fruit fly [90,91]. Moreover, comparative transcriptomic analyses have been conducted to reveal differentially expressed genes (DEGs) between B. dorsalis and its relatives [47]. These DEGs have yielded essential information concerning the molecular mechanisms of fruit fly development and adaptation, including ecdysone signaling pathways [92], cuticle biosynthesis [78], and insulin signaling [91]. Further studies have uncovered the molecular mechanisms of the B. dorsalis immune system, including peptidoglycan recognition proteins (PGRPs) [93,94], the phenoloxidase pathway [95], and toll-like receptor signaling [96,97]. Similarly, recent advancements in CRISPR-Cas9 technology have facilitated precise editing [98] of specific genes in B. dorsalis [99,100]. These findings have opened new opportunities for functional genomics research and gene-based control strategies for this insect pest. Furthermore, proteomics and metabolomics analyses have been conducted to comprehend the molecular mechanisms [101] underlying B. dorsalis growth and reproduction [102]. These studies have provided vital information on the molecular pathways and metabolites involved in insect development and adaptation. This knowledge could further assist researchers in devising innovative pest management approaches.
3.2. Reproductive Biology
Research has been conducted to understand the molecular mechanisms behind B. dorsalis mating behavior. Several neuropeptides and neurohormones involved in regulating insect reproductive behavior were identified in B. dorsalis, including corazonin, prothoracicotropic hormone, and allatostatin [103,104]. These neuropeptides play critical roles in regulating various physiological processes such as reproduction, growth, and metabolism. Furthermore, the identification of the gene encoding the alpha subunit of the cGMP-dependent protein kinase in B. dorsalis demonstrated its importance in regulating mating behavior [105,106]. Studying the receptor genes responsible for these signaling pathways would offer valuable insights into the molecular basis of B. dorsalis mating behavior. Studies focusing on the odorant receptors in B. dorsalis have been conducted. The identification of the genes encoding odorant receptors, Orco, was crucial in understanding the molecular mechanisms behind oriental fruit fly olfactory behavior and mate recognition [107,108]. Furthermore, studying the interactions between odorant receptors and their ligands, as well as the mechanisms of signal transduction in the antenna, would pave the way for developing pest management approaches in the future. The role of hormones in B. dorsalis female reproduction has been extensively investigated. Juvenile hormone (JH) plays a critical role in regulating reproduction, flight capabilities, and ecdysis in B. dorsalis. The biosynthesis, regulation, and function of JH in B. dorsalis have been thoroughly studied, and the genes responsible for JH synthesis and degradation have been cloned and characterized [86,109]. Furthermore, the 20-hydroxyecdysone (20E) pathway is another important hormonal pathway in B. dorsalis, and it has been found to play a vital role in regulating female reproduction by stimulating the biosynthesis of vitellogenin and other reproductive proteins [90,91].
Furthermore, miRNAs play a critical role in regulating B. dorsalis female reproduction through post-transcriptional regulation of target genes. Several miRNAs involved in the regulation of vitellogenin synthesis, oocyte maturation, and egg production have been identified [110,111]. The molecular mechanisms underlying the female maturation process in B. dorsalis have been studied. Critical factors in female maturation include genes involved in cuticle formation and sclerotization, ecdysteroid signaling, and ecdysone receptor signaling [87,112]. To date, the molecular mechanisms underlying the female sexual maturation process in B. dorsalis have been studied. Critical factors in female sexual maturation include genes involved in cuticle formation and sclerotization, ecdysteroid signaling, and ecdysone receptor signaling. Ongoing research on the reproductive biology and behavior of B. dorsalis is expected to yield more important findings in the near future. The discovery of additional genes and molecules involved in the regulation of B. dorsalis growth, development, and behavior could offer new strategies for developing effective means of controlling this significant pest [113].
3.3. Functional Analysis of Host Volatile Receptors
The role of volatiles in attracting B. dorsalis to crops has been well established, and partial characterization of the mechanisms of volatile detection and processing in the insect has been achieved [107,114]. However, further research is necessary to fully comprehend the complex interplay between the insect and volatile semiochemicals. This will entail a deeper exploration of the expression and function of genes involved in volatile detection, as well as identifying new semiochemicals and developing novel control strategies. The major volatile components of mango fruit that attract B. dorsalis have been identified, including hexanal, (E)-2-hexenal, (Z)-3-hexen-1-ol, and (E,E)-2,4-hexadienal [115,116]. The attractant activity of these volatiles was demonstrated through laboratory and field tests [117]. Similarly, other studies have identified attractive volatile components from other fruits, such as banana [118,119]. These findings have important implications for the development of attractive semiochemical-based control actions for B. dorsalis.
Volatile organic compounds (VOCs) from ripening fruit play a crucial role in attracting B. dorsalis. The interaction between these volatile compounds and the insect olfactory system has been extensively studied. Detection involves the diffusion of volatile compounds into the sensilla located on the antenna, which has numerous pores [120,121]. These pores facilitate the absorption of VOCs by odorant-binding proteins (OBPs) within the sensilla, which are subsequently transferred to the odorant receptors (ORs) located on the sensory neurons [122]. Using RNAi and electrophysiology techniques, researchers have been able to identify a variety of proteins involved in this process, including OBPs, chemosensory proteins (CSPs), ionotropic receptors (IRs), sensory neuron membrane proteins (SNMPs), and the odorant receptor co-receptor (ORCO) [123,124]. Silencing certain genes, such as OBPs, CSPs, and ORCO, has been shown to significantly decrease the electrophysiological response of the antennae [125,126]. In contrast to the genes primarily expressed in the antennae, there are other genes that are expressed both in classical olfactory and non-olfactory organs, or only in non-olfactory organs like the head, legs, and abdomen [127]. Further research is needed to better understand the role of these genes in attracting the oriental fruit fly to ripe fruit, which holds promise as a fruitful area for future study. As the interaction between B. dorsalis and the volatile compounds from ripening fruit is a complex and well-studied process, further research into the specific genes and proteins involved could improve our understanding of how insects are attracted to certain fruits and potentially lead to more effective agricultural pest control.
3.4. Gut Microbiota
The gut microbiota of insects is not only important for the health and survival of insects but also has the potential to impact the environment and human health [128]. Recent studies have shed light on the gut bacterial community of B. dorsalis and the role they play in insect fitness [129,130]. A study revealed that adult oriental fruit flies harbor a stable gut bacterial community dominated by Enterobacteriaceae, Klebsiella, Citrobacter, Enterobacter, and Pectobacterium [131,132]. Bacterial diversity was found to be influenced by the type of food. These gut bacteria are believed to indirectly contribute to host fitness by preventing the establishment or proliferation of pathogenic bacteria. Further investigations into the role of the Duox gene in regulating the gut bacterial community homeostasis of B. dorsalis [133,134] revealed that the suppression of the Duox gene leads to an increased bacterial load and a decreased relative abundance of Enterobacteriaceae and Leuconostocaceae in the gut. Hence, Duox plays a key role in maintaining the stability of the gut microbiome. Recent studies have demonstrated the significance of gut microbiota in the life history of B. dorsalis [135,136]. Research has shown that these bacteria play a crucial role in regulating the gut homeostasis of B. dorsalis, preventing the establishment and proliferation of pathogenic bacteria [133,137]. Some bacteria, Bacillus cereus and Enterococcus faecalis, are able to secrete volatile compounds that are capable of attracting fruit flies, suggesting that these bacteria may be potential biocontrol agents, i.e., microbial attraction baits [138].
Research on the reproductive system of female B. dorsalis found Enterobacter sakazakii and Klebsiella oxytoca as the dominating bacterial species [139]. Culture-dependent (involving isolation and microbial culture) and culture-independent techniques (using molecular techniques—without cultivation) have also been used to survey the gut and reproductive tract of B. dorsalis, revealing diverse bacterial communities in fruit flies [140]. These communities are dominated by Enterobacteriaceae, Klebsiella, Citrobacter, Enterobacter, and Pectobacterium [141]. These findings provide new insights into the role of gut microbiota in insect reproduction. In the reproductive system of female B. dorsalis, Enterobacter sakazakii and Klebsiella oxytoca are the dominant bacterial species [139,142]. These findings provide new insights into the complex interactions between the oriental fruit fly and its associated bacteria, highlighting their importance in the biology of the insect [143]. Further research is needed to better understand the mechanisms underlying these interactions and their role in the survival and fitness of B. dorsalis.
3.5. Detection and Infestation of B. dorsalis
B. dorsalis has the potential to expand and infest the northern and southern regions, making it a significant quarantine pest in China [8]. To effectively manage this pest, a comprehensive approach, including quantitative risk assessment, is necessary. Given its ability to spread through the transportation of infested fruit, strict quarantine measures must be implemented to prevent its spread to new areas. Detection of B. dorsalis can be accomplished through a combination of morphological identification, acoustic detection, and molecular detection [125,144]. To eliminate infestations, thermal treatment of infected fruit is recommended. Research has shown that treating fruit at 60 °C for 2 h or 45 °C for 5 h effectively kills all fruit flies inside [145,146], and this technique can be implemented at quarantine check points to treat infested fruit. The fly can cause significant damage to a wide range of fruit crops, leading to substantial economic losses for farmers. Its rapid spread has been attributed to its ability to adapt to new environments, high reproductive potential, and resistance to chemical insecticides [147,148]. Strict quarantine measures have been implemented in China to prevent the spread of B. dorsalis, including surveillance programs to detect the fly’s presence and thermal treatment systems to eliminate fruit infestations. In one study, B. dorsalis larvae in navel oranges were exposed to 1.2 °C for 15 days, which resulted in 99.99% larval mortality, hence making it one of the best commercial quarantine treatments [149]. Similarly, the effectiveness of other treatment measures has been studied in recent literature articles [4,8]. Moreover, as a significant quarantine pest in China, B. dorsalis requires comprehensive management strategies to control its spread. Quarantine measures, implementation of surveillance programs, and cold and thermal treatment of infested fruit have shown promising results in controlling the spread of B. dorsalis [150,151].
4. Pest Management
4.1. Mass Trapping
B. dorsalis could be mass-trapped using pheromone and food-based baits. Various pheromones and scent-based compounds, including synthetic para-pheromones or male lures such as methyl eugenol (ME), have been developed to attract and control B. dorsalis. These compounds mimic the natural pheromones produced by melon and oriental fruit flies. These pheromones are used to attract and trap male flies, allowing for population monitoring and infestation assessment [152]. E-coniferyl alcohol (E-CF) has also been found to be effective in attracting female B. dorsalis [69]. A comprehensive investigation on current resistance and lure tolerance to fruit flies [153] assessed the response of B. dorsalis males to non-ME lures. The experiment evaluated the mating and lure response of non-ME-responding (NMR) and non-responding lines (NRLs) of B. dorsalis males. Results showed that NMR males had higher mating success rates compared to NRL males and exhibited a greater attraction to non-ME lures, which have been implicated in the development of tolerance mechanisms among B. dorsalis populations [153,154].
Another research revealed that B. dorsalis causes significant economic losses in the fruit and vegetable industry by laying eggs inside hosts. Chemical controls are not very effective due to the pest’s cryptic feeding habits, strong flight ability, and resistance to insecticides. Olfaction-based trapping using ME has been the most cost-effective tool for monitoring and controlling B. dorsalis populations for seven decades [69,123]. However, laboratory selection for ME responsiveness has resulted in the non-responsiveness of B. dorsalis, which may lead to the recolonization of the pest in some areas [155]. The study aimed to determine the levels of ME responsiveness in B. dorsalis field populations in China [153,154]. Results showed that the field populations had lower ME sensitivity compared to the susceptible strain, possibly due to odorant binding protein (BdorOBP2, BdorOBP83b), and P450 gene expressions in olfactory organs [154]. Protein-based baits and food odors, such as yeast, vinegar, and fermentation products, can also attract both male and female oriental fruit flies. These baits can be combined with pheromones to increase trap efficacy [156]. Visual attractants, such as brightly colored sticky traps, can also be used to attract oriental fruit flies, and they can complement other attractants for a more comprehensive monitoring and control approach [114,157]. It is important to note that attractants can be species-specific, and the most effective ones for B. dorsalis may vary based on environmental conditions and other factors. Following are the steps involved in bait-based physical control techniques for managing B. dorsalis infestations: (a) Monitoring: It includes observations and record-keeping of the presence, distribution, and abundance of B. dorsalis in affected areas. (b) Selection of bait material: It includes selection of appropriate bait material, such as food-based baits (fishmeal or yeast hydrolysates, ME, raspberry ketone, cue lure, honey, or molasses) that have been successful in attracting fruit fly species. (c) Formulation of bait: It includes formulating the selected bait material into an attractive and easily dispersible form by adding a food-grade preservative for shelf-life extension and a hydroscopic agent to maintain its moisture content. (d) Deployment of baits: It involves deployment of the baits using various methods, including bait stations, bait trees, or spray applications, depending on the specific circumstances of each situation. (e) Collection and disposal of captured fruit flies: It involves regular monitoring to assess the effectiveness of the bait and removing and disposing of captured fruit flies to prevent escape and further spread. (f) Evaluation: It includes assessing the success of the bait-based physical control technique by monitoring oriental fruit fly population levels over time and comparing pre- and post-treatment populations to determine the reduction in the number of fruit flies.
4.2. Biological Control
Parasitoids, hymenopteran wasps, lay their eggs inside hosts, consuming them from the inside and leading to their death. Fopius arisanus (Sonan), a species of egg parasitoid, targets B. dorsalis [8]. As a potential biological control agent, F. arisanus effectively parasitizes the host eggs and reduces the pest population [158]. It is well-adapted to tropical and subtropical environments, distributed throughout Asia, Africa, and the Pacific region [159]. Utilizing F. arisanus offers advantages over chemical pest control, including specificity to the target pest, conservation of beneficial insects, and long-term sustainability [160]. In order to effectively utilize F. arisanus for biological control, it is important to understand its biology, behavior, and life cycle, as well as its interactions with the host and other factors that may affect its efficacy. Researchers have also developed mass rearing for F. arisanus to produce large numbers of individuals for release into the field. F. arisanus is a promising biological control agent for the oriental fruit fly, offering a sustainable and environmentally friendly approach to managing this destructive pest [161]. Another parasitoid, Spalangia endius (Walker) (Hymenoptera: Pteromalidae) is a solitary endoparasitoid that attacks fruit fly pupae, including B. dorsalis. This wasp lays its eggs inside the pupae, and the emerging larvae consume the host pupae from within, killing the fruit fly [162]. Using S. endius for the biological control of B. dorsalis has advantages over other methods. It is highly specific in targeting fruit fly pests and does not harm beneficial insects. Field trials have shown that this parasitoid can effectively reduce the number of B. dorsalis adults, thereby minimizing crop damage [163,164]. To effectively use S. endius for biological control, understanding the biology and behavior of both the wasp and the fruit fly is crucial. The timing of wasp releases is critical in achieving maximum parasitism rates. In general, releases should coincide with the emergence of fruit fly pupae, which is the stage at which S. endius lays its eggs. Releasing large numbers of parasitoids can help control fruit fly populations in a targeted area [165,166] (Table 1). Viruses, bacteria, and fungi can also infect and be lethal to the fruit fly adult and larvae [137]. Among the pathogens studied for use against B. dorsalis, viruses, especially baculoviruses, have been found to be highly virulent to fruit fly species. They have demonstrated effectiveness in reducing fruit fly populations in both laboratory and field studies.
Baculoviruses are insect-specific viruses that replicate within the insect host and cause death. Among the baculovirus isolates identified and characterized, the nuclear polyhedrosis virus (NPV) has been found to be highly virulent to several insect species [167,168]. NPV studies have demonstrated its ability to reduce the number of fruit fly individuals in laboratory and field settings, thereby decreasing the damage caused by this pest. Moreover, NPV is safe for the environment and non-target organisms, making it a promising option for fruit fly management [169]. However, it is important to note that using pathogenic microorganisms, including viruses, for insect pest management is still in its early stages, and more research is needed to fully understand their potential and limitations. Baculoviruses, particularly NPV, have shown potential for controlling B. dorsalis, and further research is needed to integrate them into pest management programs effectively. The entomophagous fungus Beauveria bassiana (Sordariomycetes: Clavicipitaceae) is an entomopathogenic fungus that is known to be an effective biological control agent against B. cucurbitae. This fungus infects the insects and causes mortality [170]. In China, B. bassiana effectively controlled B. dorsalis, achieving a mortality rate of over 80% in laboratory experiments. Similarly, another study showed that B. bassiana effectively reduced the population density of B. dorsalis in the field [171,172]. B. bassiana can be used as a biological control agent in several ways: (1) Inoculative releases: This involves releasing large numbers of fungal spores (conidia) into the environment, which then infect the insects. This approach is most effective when used in conjunction with other management strategies, such as the use of pheromones or host-plant resistance. (2) Injection or spraying: This process involves injecting or spraying a suspension of conidia directly onto the insects, causing them to become infected. (3) Formulations: B. bassiana can also be formulated into granules or dusts that can be applied to the host plants or environment, where they will encounter the insects. The entomopathogenic bacterium Bacillus thuringiensis (Bt) is a naturally occurring soil bacterium that produces a toxic crystal protein effective against many insect pests, including B. dorsalis [173]. Entomopathogenic nematodes are parasitic roundworms that can infect and kill fruit fly larvae [174]. Further research and understanding of biology will improve their integration into pest management programs (Table 1).

{kind=link}
{kind=link}
{kind=link}
Table 1.
Natural enemies of B. dorsalis.
| Bio-Control Agents | Name of Species | Host Stages | Reference |
|---|---|---|---|
| Predator | Oecophylla longinoda | Pupa/larva | [175] |
| Pachycrepoideus vindemmiae | Larva/pupa | [176] | |
| Parasitoids | Fopius arisanus | Egg | [177] |
| Psyttalia cosyrae | Larva-pupal | [178] | |
| Diachasmimorpha longicaudata | Larva | [178] | |
| Nematodes | Heterorhabditis taysearae | Larva/pupa | [176] |
| H. indica | Larva/pupa | [179] | |
| Steinernema sp | Larva/pupa | [180] |
4.3. Sterile Insect Technique (SIT)
The sterile insect technique is a promising biological control for Bactrocera species, with a proven track record of success in various countries worldwide. It is a sustainable and eco-friendly method of pest management that complements other control strategies, providing long-term control of this economically important insect pest. The technique has been used for decades to manage various insect pests, including B. dorsalis, commonly known as the oriental fruit fly. SIT involves mass-rearing and sterilization of male insects, which are then released into the wild to mate with females. Mating with sterilized males leads to the laying of eggs by female insects that do not hatch, ultimately leading to a decline in the pest population [8,181]. This technique was first employed in the 1950s to control the screwworm, Cochliomyia hominivorax, in the southern United States and has since been effectively utilized against various other insect pests worldwide. Fruit flies, including B. dorsalis, have been successfully managed using SIT in several countries, such as China, Australia, and Hawaii [182,183]. For instance, in Hawaii, SIT was implemented in the early 2000s to manage oriental fruit fly outbreaks in the state’s agriculture industry. The program’s success resulted in a significant decline in the pest population [184]. In Australia, SIT has been incorporated into integrated pest management to control Mediterranean fruit fly C. capitata populations in the country’s horticulture industry [185]. The genetic sexing strain is a technique utilized to manipulate the sex ratios of a population, leading to more effective and efficient pest management. It has been successfully applied worldwide, including China, to manage B. dorsalis. This technique employs a genetic marker to distinguish between male and female fruit flies. By releasing only sterilized males into the environment, the population growth of the pest can be suppressed without the need for chemical insecticides [186].
In China, researchers have developed a genetic sexing strain for B. dorsalis using the temperature-sensitive lethal (tsl) mutation. This mutation causes the death of females at a certain temperature, enabling the separation of male and female fruit flies [187]. The genetic sexing strain has been proven effective in suppressing the population growth of several fruit fly species in field trials [188]. This technique has also been applied to manage fruit flies in other countries, including Australia and Thailand [13,185,189,190]. These studies demonstrate the potential of the genetic sexing strain as an integrated pest management tool for managing tephritid fruit flies.
4.4. Molecular Control
The management of B. dorsalis is challenging due to its high resistance to insecticides. To overcome this challenge, it is crucial to identify new targets for insect pest control. Transient receptor potential (TRP) channels play a crucial role in various physiological processes in insects, including nociception, thermo-sensation, and olfaction [191,192]. In recent years, there have been extensive studies on the identification and characterization of TRP channels in various insect species, including B. dorsalis. In one study [192], 15 TRP channel genes were identified in the genome of B. dorsalis. The expression patterns of these genes were analyzed in different tissues, such as the antennae, brain, midgut, Malpighian tubules, and fat body. The results revealed that TRP channels were differentially expressed across various tissues, with some TRP genes being predominantly expressed in specific tissues. Additionally, another study [193] investigated the role of TRP channels in insecticide resistance in insects. They used RNA interference (RNAi) to knock down the expression of TRP channels in B. dorsalis. The findings showed that knockdown of TRP channels significantly reduced insecticide resistance in B. dorsalis, suggesting the potential utilization of TRP channels as targets for insect pest control [194,195]. The identification, characterization, and expression analysis of TRP channel genes in the oriental fruit fly will provide crucial information for the development of new and effective strategies for the management and control of this pest.
4.5. RNA Interference (RNAi)
RNA interference (RNAi) is a highly effective technique for gene silencing through the use of double-stranded RNA (dsRNA) [196]. It has shown promise in knocking down insect pests as a more environmentally friendly option. Previous studies have demonstrated successful silencing of genes rpl19, v-ATPase-D, noa, and rab11 in adult B. dorsalis through the feeding of corresponding dsRNA. Other potential target genes involved in midgut digestion and detoxification have also been identified [197,198]. However, using RNAi for controlling the oriental fruit fly faces challenges, including effectively delivering dsRNA to the insect and potential risks to non-target organisms. The delivery of dsRNA has not been fully implemented yet, and the possible impacts on non-target organisms and host fruits and vegetables must be carefully considered. There is a risk of reducing the expression of genes in natural enemies and other beneficial insects due to the high similarity in rpl19 sequences between these insects and B. dorsalis. Therefore, minimizing the impact of dsRNA on non-target insects and host fruits and vegetables is a priority in ongoing efforts to use RNAi for controlling B. dorsalis. In a research article addressing the problem of insecticide resistance in B. dorsalis, a global pest affecting various crops, researchers focused on the role of UDP-glycosyltransferases (UGTs) in resistance development [199]. These enzymes are involved in metabolically processing both plant secondary metabolites and synthetic insecticides. The study identified 31 UGT genes in the genome of B. dorsalis, with 12 of them highly expressed in key tissues such as the antennae, midgut, Malpighian tubules, and fat body. Furthermore, exposure to four different insecticides caused a significant upregulation of 17 UGT genes. To investigate further, RNA interference was used to knock down five selected UGT genes, resulting in reduced oriental fruit fly mortality in response to insecticides from 9.29% to 27.22% [200].
4.6. CRISPR-Cas9
The clustered regularly interspaced palindromic repeat (CRISPR-Cas9) system is a revolutionary tool for precise and efficient genome editing in various organisms [201]. In a study of B. dorsalis, researchers targeted a specific gene known as the Sex Peptide Receptor (Bdspr) using CRISPR/Cas9 technology [100]. The Bdspr gene plays a critical role in the regulation of female reproduction, including ovary development and egg laying. By introducing mutations into this gene, the researchers aimed to examine its effects on female fecundity and reproductive functions in B. dorsalis. Several research experiments showed that CRISPR/Cas9-mediated disruption of the Bdspr gene, when the insects were fed with the ds-spr gene, led to significant changes in the number and size of ovarioles, a reduction in the number of eggs laid, and a decrease in overall female fecundity. This indicated the importance of the Bdspr gene in the normal functioning of the female reproductive system in B. dorsalis. The study also demonstrated that the CRISPR/Cas9 system is an effective tool for studying gene function and disrupting specific genes in insects. In the future, this information could potentially be used to develop new strategies for controlling the population of oriental fruit flies, a major agricultural pest causing significant damage to crops worldwide [100,202,203,204]. The CRISPR/Cas9-induced mutation of the Bdspr gene in the oriental fruit fly underscores the significance of this gene in female reproduction and highlights the potential of genome editing technology for advancing the field of insect pest management.
In another study focused on understanding the functional role of the white gene in pigmentation in B. dorsalis, the white gene was cloned, and knockout strains were created using the CRISPR/Cas9 genome editing system. The results revealed that the mutants lost pigmentation in the compound eye and their head spots. Further analysis using quantitative reverse-transcription PCR showed lower expression levels of the Bd-yellow1 gene in the head of mutants compared to the wild-type strain, while there were no significant differences in the expression of the other six genes. As the yellow gene is crucial for melanin biosynthesis, the reduced expression of Bd-yellow1 in mutants led to a decrease in dark pigmentation in the head spots. This study provides evidence for the first time that the white gene may play a role in cuticle pigmentation by affecting the expression of the yellow gene [99].
5. Conclusions and Future Perspective
The oriental fruit fly has been the most common and significant orchard pest since its invasion of mainland China. Currently, it is expanding to more suitable regions in North China. Over the past few decades, many tactics have been developed to track its occurrence, spread, and damage, along with numerous studies conducted to understand its invasion process. However, its ability to adapt to different habitats, insecticide resistance, and high reproductive capacity have been helping the pest to spread to wider landscapes. These factors might provide valuable information for the development of new pest management techniques. Currently, the application of synthetic pesticides remains the basis of B. dorsalis management worldwide, including China. However, when this pest develops insecticide resistance, the effectiveness of chemical management significantly reduces. To address this issue, it is essential to take preventive measures against the emergence of insecticide resistance and adopt novel pesticide options, such as botanical and microbial pesticides. Gaining a sufficient understanding of the physiological mechanism and molecular roots of mating choice and behavior could lead to new control techniques based on behavioral alteration. SIT (sterile insect technique) and area-wide pest eradication programs have been used as techniques to manage the oriental fruit fly and other insect species. Although it has been widely used in China, it has not yet eradicated the invasive species. However, it holds the potential to successfully eradicate the pest species.
Author Contributions
Conceptualization, S.J. and Y.L.; literature search, S.J.; software, S.J. and S.A.H.R.; validation, Y.L. and S.J.; formal analysis, S.J.; investigation, Y.L.; resources, Y.L.; data curation, S.J.; writing—original draft preparation, S.J.; writing—review and editing, S.J., Y.L.; visualization, Y.L.; funding acquisition, Y.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Common Key Technology R & D Innovation Team Project in Modern Agricultural Industry of Guangdong grant number 2022kj134.
Data Availability Statement
The data used in this study will be made available by the corresponding author on request.
Acknowledgments
The authors thank all anonymous reviewers for their constructive comments, invaluable suggestions, and recommendations.
Conflicts of Interest
The authors declare they have no conflicts of interest.
References
- He, Y.; Xu, Y.; Chen, X. Biology, Ecology and Management of Tephritid Fruit Flies in China: A Review. Insects 2023, 14, 196. [Google Scholar] [CrossRef] [PubMed]
- Clarke, A.R.; Armstrong, K.F.; Carmichael, A.E.; Milne, J.R.; Raghu, S.; Roderick, G.K.; Yeates, D.K. Invasive phytophagous pests arising through a recent tropical evolutionary radiation: The Bactrocera dorsalis complex of fruit flies. Annu. Rev. Entomol. 2005, 50, 293–319. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.; Sarkar, B.; Hossain, M.; Mian, M.; Rajotte, E.; Muniappan, R.; O’Rourke, M. Comparison of biorational management approaches against mango fruit fly (Bactrocera dorsalis Hendel) in Bangladesh. Crop Prot. 2020, 135, 104807. [Google Scholar] [CrossRef]
- Jaffar, S.; Lu, Y. Toxicity of Some Essential Oils Constituents against Oriental Fruit Fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Insects 2022, 13, 954. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.; Liu, Y.; Zhang, B. Invasion history of the oriental fruit fly, Bactrocera dorsalis, in the Pacific-Asia region: Two main invasion routes. PLoS ONE 2012, 7, e36176. [Google Scholar] [CrossRef] [PubMed]
- Drew, R.; Hancock, D. Biogeography, Speciation and Taxonomy within the genus Bactrocera Macquart with application to the Bactrocera dorsalis (Hendel) complex of fruit flies (Diptera: Tephritidae: Dacinae). Zootaxa 2022, 5190, 333–360. [Google Scholar] [CrossRef]
- Zeng, Y.; Reddy, G.V.; Li, Z.; Qin, Y.; Wang, Y.; Pan, X.; Jiang, F.; Gao, F.; Zhao, Z.H. Global distribution and invasion pattern of oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae). J. Appl. Entomol. 2019, 143, 165–176. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, D.; Xu, Y.; Wang, L.; Cheng, D.; Qi, Y.; Zeng, L.; Lu, Y. Invasion, expansion, and control of Bactrocera dorsalis (Hendel) in China. J. Integr. Agric. 2019, 18, 771–787. [Google Scholar] [CrossRef]
- Aketarawong, N.; Guglielmino, C.; Karam, N.; Falchetto, M.; Manni, M.; Scolari, F.; Gomulski, L.; Gasperi, G.; Malacrida, A. The oriental fruitfly Bactrocera dorsalis ss in East Asia: Disentangling the different forces promoting the invasion and shaping the genetic make-up of populations. Genetica 2014, 142, 201–213. [Google Scholar] [CrossRef]
- Meng, L.W.; Yuan, G.R.; Lu, X.P.; Jing, T.X.; Zheng, L.S.; Yong, H.X.; Wang, J.J. Two delta class glutathione S-transferases involved in the detoxification of malathion in Bactrocera dorsalis (Hendel). Pest. Manag. Sci. 2019, 75, 1527–1538. [Google Scholar] [CrossRef]
- Mutamiswa, R.; Nyamukondiwa, C.; Chikowore, G.; Chidawanyika, F. Overview of oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae) in Africa: From invasion, bio-ecology to sustainable management. Crop Prot. 2021, 141, 105492. [Google Scholar] [CrossRef]
- Zhu, Y.; Qi, F.; Tan, X.; Zhang, T.; Teng, Z.; Fan, Y.; Wan, F.; Zhou, H. Use of Age-Stage, Two-Sex Life Table to Compare the Fitness of Bactrocera dorsalis (Diptera: Tephritidae) on Northern and Southern Host Fruits in China. Insects 2022, 13, 258. [Google Scholar] [CrossRef] [PubMed]
- Orankanok, W.; Chinvinijkul, S.; Thanaphum, S.; Sitilob, P.; Enkerlin, W.R. Area-Wide Integrated Control of Oriental Fruit Fly Bactrocera dorsalis and Guava Fruit Fly Bactrocera correcta in Thailand. In Area-Wide Control of Insect Pests; Springer: Dordrecht, The Netherlands, 2007; pp. 517–526. [Google Scholar]
- Bana, J.; Sharma, H.; Kumar, S.; Singh, P. Impact of weather parameters on population dynamics of oriental fruit fly, Bactrocera dorsalis (Hendel)(Diptera: Tephritidae) under south Gujarat mango ecosystem. J. Agrometeorol. 2017, 19, 78–80. [Google Scholar] [CrossRef]
- Jitendra, M.; Sandeep, S.; Akilesh, T.; Chaube, M. Population dynamics of oriental fruit fly, Bactrocera dorsalis (Hendel) in relation to abiotic factors. HortFlora Res. Spectr. 2012, 1, 187–189. [Google Scholar]
- Dhillon, M.; Singh, R.; Naresh, J.; Sharma, H. The melon fruit fly, Bactrocera cucurbitae: A review of its biology and management. J. Insect Sci. 2005, 5, 40. [Google Scholar] [CrossRef]
- Coates, B.S.; Poelchau, M.; Childers, C.; Evans, J.D.; Handler, A.; Guerrero, F.; Skoda, S.; Hopper, K.; Wintermantel, W.M.; Ling, K.-S. Arthropod genomics research in the United States Department of Agriculture-Agricultural Research Service: Current impacts and future prospects. Trends Entomol. 2015, 11, 12–27. [Google Scholar]
- Liu, L.J.; Martinez-Sañudo, I.; Mazzon, L.; Prabhakar, C.S.; Girolami, V.; Deng, Y.L.; Dai, Y.; Li, Z.H. Bacterial communities associated with invasive populations of Bactrocera dorsalis (Diptera: Tephritidae) in China. Bull. Entomol. Res. 2016, 106, 718–728. [Google Scholar] [CrossRef]
- Zhu, Y.-F.; Tan, X.-M.; Qi, F.-J.; Teng, Z.-W.; Fan, Y.-J.; Shang, M.-Q.; Lu, Z.-Z.; Wan, F.-H.; Zhou, H.-X. The host shift of Bactrocera dorsalis: Early warning of the risk of damage to the fruit industry in northern China. Entomol. Gen. 2022, 42, 691–699. [Google Scholar] [CrossRef]
- Ullah, F.; Gul, H.; Hafeez, M.; Güncan, A.; Tariq, K.; Desneux, N.; Zhao, Z.; Li, Z. Impact of temperature stress on demographic traits and population projection of Bactrocera dorsalis. Entomol. Gen. 2022, 42, 949–957. [Google Scholar] [CrossRef]
- Doorenweerd, C.; Leblanc, L.; Hsu, Y.-F.; Huang, C.-L.; Lin, Y.-C.; San Jose, M.; Rubinoff, D. Taiwan’s Dacini Fruit Flies: Rare Endemics and Abundant Pests, along Altitudinal Gradients. Pac. Sci. 2019, 73, 35–59. [Google Scholar] [CrossRef]
- Jing, W.; QI, G.-j.; Jun, M.; Ren, Y.; Rui, W.; McKirdy, S. Predicting the potential geographic distribution of Bactrocera bryoniae and Bactrocera neohumeralis (Diptera: Tephritidae) in China using MaxEnt ecological niche modeling. J. Integr. Agric. 2020, 19, 2072–2082. [Google Scholar]
- Li, X.; Yang, H.; Wang, T.; Wang, J.; Wei, H. Life history and adult dynamics of Bactrocera dorsalis in the citrus orchard of Nanchang, a subtropical area from China: Implications for a control timeline. Scienceasia 2019, 45, 212–220. [Google Scholar] [CrossRef]
- Qin, Y.; Wang, C.; Zhao, Z.; Pan, X.; Li, Z. Climate change impacts on the global potential geographical distribution of the agricultural invasive pest, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Clim. Chang. 2019, 155, 145–156. [Google Scholar] [CrossRef]
- Kim, S.B.; Park, J.-J.; Kim, D.-S. CLIMEX simulated predictions of the potential distribution of Bactrocera dorsalis (Hendel) (Diptera: Tephritidae) considering the northern boundary: With special emphasis on Jeju, Korea. J. Asia-Pac. Entomol. 2020, 23, 797–808. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, S.; De Meyer, M.; Liao, Z.; Zhao, Y.; Virgilio, M.; Feng, S.; Qin, Y.; Singh, S.; Wee, S.L. Genomes of the cosmopolitan fruit pest Bactrocera dorsalis (Diptera: Tephritidae) reveal its global invasion history and thermal adaptation. J. Adv. Res. 2022. [Google Scholar] [CrossRef]
- Kim, H.; Kim, S.; Kim, S.; Lee, Y.; Lee, H.-S.; Lee, S.-J.; Choi, D.-S.; Jeon, J.; Lee, J.-H. Population genetics for inferring introduction sources of the oriental fruit fly, Bactrocera dorsalis: A test for quarantine use in Korea. Insects 2021, 12, 851. [Google Scholar] [CrossRef]
- Ouyang, H.; Wang, X.; Zheng, X.; Lu, W.; Qin, F.; Chen, C. Full-length SMRT transcriptome sequencing and SSR analysis of Bactrocera dorsalis (Hendel). Insects 2021, 12, 938. [Google Scholar] [CrossRef]
- Li, Y.; Wu, Y.; Chen, H.; Wu, J.; Li, Z. Population structure and colonization of Bactrocera dorsalis (Diptera: Tephritidae) in China, inferred from mtDNA COI sequences. J. Appl. Entomol. 2012, 136, 241–251. [Google Scholar] [CrossRef]
- Liu, J.; Shi, W.; Ye, H. Population genetics analysis of the origin of the Oriental fruit fly, Bactrocera dorsalis Hendel (Diptera: Tephritidae), in northern Yunnan Province, China. Entomol. Sci. 2007, 10, 11–19. [Google Scholar] [CrossRef]
- Moschilla, J.A.; Tomkins, J.L.; Simmons, L.W. Males adjust their manipulation of female remating in response to sperm competition risk. Proc. Biol. Sci. 2020, 287, 20201238. [Google Scholar] [CrossRef]
- Wei, D.; Feng, Y.-C.; Wei, D.-D.; Yuan, G.-R.; Dou, W.; Wang, J.-J. Female remating inhibition and fitness of Bactrocera dorsalis (Diptera: Tephritidae) associated with male accessory glands. Fla. Entomol. 2015, 98, 52–58. [Google Scholar] [CrossRef]
- Reyes-Hernández, M.; Thimmappa, R.; Abraham, S.; Pagadala Damodaram, K.J.; Pérez-Staples, D. Methyl eugenol effects on Bactrocera dorsalis male total body protein, reproductive organs and ejaculate. J. Appl. Entomol. 2019, 143, 177–186. [Google Scholar] [CrossRef]
- Shelly, T.E. Fecundity of Female Oriental Fruit Flies (Diptera: Tephritidae): Effects of Methyl Eugenol-Fed and Multiple Mates. Ann. Entomol. Soc. Am. 2000, 93, 559–564. [Google Scholar] [CrossRef]
- Shelly, T. Ability of sterile males to inhibit female remating in the oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Proc. Hawaii. Entomol. Soc. 2020, 52, 15–23. [Google Scholar]
- Olabiyi, D.; Shrestha, B.; Zaka, S.M.; Neupane, S. Insect Pests of Citrus Production. In Citrus Production; CRC Press: Boca Raton, FL, USA, 2022; pp. 287–302. [Google Scholar]
- Kota, M.; Olzer, R. Insect Communication. In Encyclopedia of Animal Cognition and Behavior; Vonk, J., Shackelford, T., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–7. [Google Scholar] [CrossRef]
- Meunier, J. Social immunity and the evolution of group living in insects. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140102. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, Y.; Yang, L.; Chen, W.; Jiang, Z.; Xiao, Z.; Xie, X.; Zhong, G.; Yi, X. Group housing enhances mating and increases the sensitization of chemical cues in Bactrocera dorsalis. Pest. Manag. Sci. 2023, 79, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Kamala Jayanthi, P.D.; Saravan Kumar, P.; Vyas, M. Odour Cues from Fruit Arils of Artocarpus heterophyllus Attract both Sexes of Oriental Fruit Flies. J. Chem. Ecol. 2021, 47, 552–563. [Google Scholar] [CrossRef]
- Wang, L.; Anderson, D.J. Identification of an aggression-promoting pheromone and its receptor neurons in Drosophila. Nature 2010, 463, 227–231. [Google Scholar] [CrossRef]
- Billeter, J.-C.; Levine, J. The role of cVA and the Odorant binding protein Lush in social and sexual behavior in Drosophila melanogaster. Front. Ecol. Evol. 2015, 3, 75. [Google Scholar] [CrossRef]
- Ejima, A.; Smith, B.P.; Lucas, C.; Van Naters, W.v.d.G.; Miller, C.J.; Carlson, J.R.; Levine, J.D.; Griffith, L.C. Generalization of courtship learning in Drosophila is mediated by cis-vaccenyl acetate. Curr. Biol. 2007, 17, 599–605. [Google Scholar] [CrossRef]
- Roy, A.; George, S.; Palli, S.R. Multiple functions of CREB-binding protein during postembryonic development: Identification of target genes. BMC Genom. 2017, 18, 996. [Google Scholar] [CrossRef]
- Geib, S.M.; Calla, B.; Hall, B.; Hou, S.; Manoukis, N.C. Characterizing the developmental transcriptome of the oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae) through comparative genomic analysis with Drosophila melanogaster utilizing modENCODE datasets. BMC Genom. 2014, 15, 942. [Google Scholar] [CrossRef]
- De Villiers, M.; Hattingh, V.; Kriticos, D.J.; Brunel, S.; Vayssières, J.F.; Sinzogan, A.; Billah, M.K.; Mohamed, S.A.; Mwatawala, M.; Abdelgader, H.; et al. The potential distribution of Bactrocera dorsalis: Considering phenology and irrigation patterns. Bull. Entomol. Res. 2016, 106, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Zhao, Y.; Su, Y.; Wu, J.; Wang, Z.; Hu, J.; Liu, L.; Zhao, Z.; Hoffmann, A.A.; Chen, B. A transcriptional and functional analysis of heat hardening in two invasive fruit fly species, Bactrocera dorsalis and Bactrocera correcta. Evol. Appl. 2019, 12, 1147–1163. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Lu, Y.-Y.; Zeng, L.; Pang, S.-T. Effect of different hostfruits on the cold hardiness of Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Acta Entomol. Sin. 2006, 49, 447–453. [Google Scholar]
- Mutamiswa, R.; Tarusikirwa, V.; Nyamukondiwa, C.; Chidawanyika, F. Fluctuating environments impact thermal tolerance in an invasive insect species Bactrocera dorsalis (Diptera: Tephritidae). J. Appl. Entomol. 2020, 144, 885–896. [Google Scholar] [CrossRef]
- Pieterse, W.; Terblanche, J.S.; Addison, P. Do thermal tolerances and rapid thermal responses contribute to the invasion potential of Bactrocera dorsalis (Diptera: Tephritidae)? J. Insect Physiol. 2017, 98, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Jia, F.-X.; Dou, W.; Hu, F.; Wang, J.-J. Effects of thermal stress on lipid peroxidation and antioxidant enzyme activities of oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae). Fla. Entomol. 2011, 94, 956–963. [Google Scholar] [CrossRef]
- Gao, X.M.; Jia, F.X.; Shen, G.M.; Jiang, H.Q.; Dou, W.; Wang, J.J. Involvement of superoxide dismutase in oxidative stress in the oriental fruit fly, Bactrocera dorsalis: Molecular cloning and expression profiles. Pest. Manag. Sci. 2013, 69, 1315–1325. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Dou, W.; Jiang, M.; Wang, J. Oriental Fruit Fly Bactrocera dorsalis (Hendel). In Biological Invasions and Its Management in China: Volume 1; Wan, F., Jiang, M., Zhan, A., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 267–283. [Google Scholar] [CrossRef]
- Hou, B.; Xie, Q.; Zhang, R. Depth of pupation and survival of the Oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae) pupae at selected soil moistures. Appl. Entomol. Zool. 2006, 41, 515–520. [Google Scholar] [CrossRef]
- Jaffar, S.; Ahmad, S.; Lu, Y. Contribution of insect gut microbiota and their associated enzymes in insect physiology and biodegradation of pesticides. Front. Microbiol. 2022, 13, 979383. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Zeng, L.; Lin, Y.; Lu, Y.; Liang, G. Insecticide resistance of the oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae), in mainland China. Pest. Manag. Sci. 2011, 67, 370–376. [Google Scholar] [CrossRef]
- Abubakar, M.; Ali, H.; Shad, S.A.; Anees, M.; Binyameen, M. Trichlorfon resistance: Its stability and impacts on biological parameters of Bactrocera zonata (Diptera: Tephritidae). Appl. Entomol. Zool. 2021, 56, 473–482. [Google Scholar] [CrossRef]
- Lin, Y.; Jin, T.; Zeng, L.; Lu, Y. Cuticular penetration of β-cypermethrin in insecticide-susceptible and resistant strains of Bactrocera dorsalis. Pestic. Biochem. Physiol. 2012, 103, 189–193. [Google Scholar] [CrossRef]
- Wang, J.-J.; Wei, D.; Dou, W.; Hu, F.; Liu, W.-F.; Wang, J.-J. Toxicities and Synergistic Effects of Several Insecticides Against the Oriental Fruit Fly (Diptera: Tephritidae). J. Econ. Entomol. 2013, 106, 970–978. [Google Scholar] [CrossRef]
- Zhang, R.; He, S.; Chen, J. Monitoring of Bactrocera dorsalis (Diptera: Tephritidae) Resistance to Cyantraniliprole in the South of China. J. Econ. Entomol. 2014, 107, 1233–1238. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Wang, J.; Che, W.; Luo, C. First report of field resistance to cyantraniliprole, a new anthranilic diamide insecticide, on Bemisia tabaci MED in China. J. Integr. Agric. 2018, 17, 158–163. [Google Scholar] [CrossRef]
- Lu, X.P.; Wang, L.L.; Huang, Y.; Dou, W.; Chen, C.T.; Wei, D.; Wang, J.J. The epsilon glutathione S-transferases contribute to the malathion resistance in the oriental fruit fly, Bactrocera dorsalis (Hendel). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2016, 180, 40–48. [Google Scholar] [CrossRef]
- Jing, T.-X.; Wang, D.-F.; Ma, Y.-P.; Zeng, L.-L.; Meng, L.-W.; Zhang, Q.; Dou, W.; Wang, J.-J. Genome-wide and expression-profiling analyses of the cytochrome P450 genes in Bactrocera dorsalis (Hendel) and screening of candidate P450 genes associated with malathion resistance. Pest. Manag. Sci. 2020, 76, 2932–2943. [Google Scholar] [CrossRef]
- He, L.; Huang, Y.; Tang, X. RNAi-based pest control: Production, application and the fate of dsRNA. Front. Bioeng. Biotechnol. 2022, 10, 1080576. [Google Scholar] [CrossRef]
- Yan, S.; Ren, B.-Y.; Shen, J. Nanoparticle-mediated double-stranded RNA delivery system: A promising approach for sustainable pest management. Insect Sci. 2021, 28, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Hou, Q.-L.; Chen, E.-H.; Dou, W.; Wang, J.-J. Assessment of Bactrocera dorsalis (Diptera: Tephritidae) Diets on Adult Fecundity and Larval Development: Insights Into Employing the Sterile Insect Technique. J. Insect Sci. 2020, 20, 1–8. [Google Scholar] [CrossRef]
- Jing, T.X.; Zhang, Y.X.; Dou, W.; Jiang, X.Y.; Wang, J.J. First Insights into the Intrapuparial Development of Bactrocera dorsalis (Hendel): Application in Predicting Emergence Time for Tephritid Fly Control. Insects 2019, 10, 283. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, X.; Chen, Z.; Wang, Z.; Lu, Y.; Cheng, D. The Divergence in Bacterial Components Associated with Bactrocera dorsalis across Developmental Stages. Front. Microbiol. 2018, 9, 114. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, D.D.; Wan, L.; Hu, Z.Y.; He, T.T.; Wang, J.B.; Deng, S.Z.; Wang, X.S. Assessment of attractancy and safeness of (E)-coniferyl alcohol for management of female adults of Oriental fruit fly, Bactrocera dorsalis (Hendel). Pest. Manag. Sci. 2022, 78, 1018–1028. [Google Scholar] [CrossRef]
- Cheng, D.; Guo, Z.; Riegler, M.; Xi, Z.; Liang, G.; Xu, Y. Gut symbiont enhances insecticide resistance in a significant pest, the oriental fruit fly Bactrocera dorsalis (Hendel). Microbiome 2017, 5, 13. [Google Scholar] [CrossRef]
- Hu, F.; Dou, W.; Wang, J.J.; Jia, F.X.; Wang, J.J. Purification and biochemical characterization of glutathione S-transferases from four field populations of Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Arch. Insect Biochem. Physiol. 2011, 78, 201–215. [Google Scholar] [CrossRef]
- Huang, Y.-B.; Chi, H. Assessing the application of the jackknife and bootstrap techniques to the estimation of the variability of the net reproductive rate and gross reproductive rate: A case study in Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae). J. Agric. For. 2012, 61, 37–45. [Google Scholar]
- Amiri, A.; Bandani, A.R.; Kafil, M. Gut compartments and ovary bacterial symbionts of the Sunn pest. J. Asia-Pac. Entomol. 2020, 23, 723–730. [Google Scholar] [CrossRef]
- Zhou, X.; Ye, Y.-Z.; Ogihara, M.H.; Takeshima, M.; Fujinaga, D.; Liu, C.-W.; Zhu, Z.; Kataoka, H.; Bao, Y.-Y. Functional analysis of ecdysteroid biosynthetic enzymes of the rice planthopper, Nilaparvata lugens. Insect Biochem. Mol. Biol. 2020, 123, 103428. [Google Scholar] [CrossRef]
- Dhadialla, T.S.; Retnakaran, A.; Smagghe, G. 6.3—Insect Growth- and Development-Disrupting Insecticides. In Comprehensive Molecular Insect Science; Gilbert, L.I., Ed.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 55–115. [Google Scholar] [CrossRef]
- Baehrecke, E.H. Ecdysone signaling cascade and regulation of Drosophila metamorphosis. Arch. Insect Biochem. Physiol. 1996, 33, 231–244. [Google Scholar] [CrossRef]
- Uyehara, C.M.; McKay, D.J. Direct and widespread role for the nuclear receptor EcR in mediating the response to ecdysone in Drosophila. Proc. Natl. Acad. Sci. USA 2019, 116, 9893–9902. [Google Scholar] [CrossRef]
- Yang, W.-J.; Xu, K.-K.; Cong, L.; Wang, J.-J. Identification, mRNA Expression, and Functional Analysis of Chitin Synthase 1 Gene and Its Two Alternative Splicing Variants in Oriental Fruit Fly, Bactrocera dorsalis. Int. J. Biol. Sci. 2013, 9, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Fujii, S.; Kobayashi, S.; Tettey, P.A.; Lisdiana, L.; Saito, H.; Kawai, K.; Sambongi, Y. Transcriptome analysis of Chironomus sulfurosus larvae living in acidic environments: Insights into molecular mechanisms for acid tolerance. J. Insect Physiol. 2021, 133, 104288. [Google Scholar] [CrossRef] [PubMed]
- Laurie-Ahlberg, C.C.; Williamson, J.H.; Cochrane, B.J.; Wilton, A.N.; Chasalow, F.I. Autosomal factors with correlated effects on the activities of the glucose 6-phosphate and 6-phosphogluconate dehydrogenases in Drosophila melanogaster. Genetics 1981, 99, 127–150. [Google Scholar] [CrossRef]
- Yang, W.J.; Xu, K.K.; Zhang, R.Y.; Dou, W.; Wang, J.J. Transcriptional regulation of a chitinase gene by 20-hydroxyecdysone and starvation in the oriental fruit fly, Bactrocera dorsalis. Int. J. Mol. Sci. 2013, 14, 20048–20063. [Google Scholar] [CrossRef]
- Yu, L.; Zhou, Q.; Zhang, C.; Pignoni, F. Identification of Bombyx atonal and functional comparison with the Drosophila atonal proneural factor in the developing fly eye. Genesis 2012, 50, 393–403. [Google Scholar] [CrossRef]
- Bakshi, A.; Moin, M.; Madhav, M.S.; Kirti, P.B. Target of rapamycin, a master regulator of multiple signalling pathways and a potential candidate gene for crop improvement. Plant Biol. 2019, 21, 190–205. [Google Scholar] [CrossRef]
- Yoveva, A.; Sawarkar, R. Chromatin Immunoprecipitation (ChIP) of Heat Shock Protein 90 (Hsp90). Methods Mol. Biol. 2018, 1709, 221–231. [Google Scholar] [CrossRef]
- Chen, B.; Wagner, A. Hsp90 is important for fecundity, longevity, and buffering of cryptic deleterious variation in wild fly populations. BMC Evol. Biol. 2012, 12, 25. [Google Scholar] [CrossRef]
- Chen, P.; Chen, M.; Ye, H.; Yuan, R.; Du, C.; Ong, S.P. Juvenile hormone regulation on the flight capability of Bactrocera dorsalis (Diptera: Tephritidae). J. Asia-Pac. Entomol. 2020, 23, 1120–1129. [Google Scholar] [CrossRef]
- Shi, Y.; Liu, T.Y.; Jiang, H.B.; Liu, X.Q.; Dou, W.; Park, Y.; Smagghe, G.; Wang, J.J. The Ecdysis Triggering Hormone System, via ETH/ETHR-B, Is Essential for Successful Reproduction of a Major Pest Insect, Bactrocera dorsalis (Hendel). Front. Physiol. 2019, 10, 151. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-H.; Xia, Y.-D.; Zhang, Q.; Li, W.; Li, R.-Y.; Liu, Y.; Chen, E.-H.; Dou, W.; Stelinski, L.L.; Wang, J.-J. Potential targets for controlling Bactrocera dorsalis using cuticle- and hormone-related genes revealed by a developmental transcriptome analysis. Pest. Manag. Sci. 2020, 76, 2127–2143. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, S.; Srivastava, P.; Rathore, D.P.; Pandey, A.K. Amylases: A Prospective Enzyme in the Field of Biotechnology. J. Appl. Biosci. 2015, 41, 1–18. [Google Scholar]
- Ekoka, E.; Maharaj, S.; Nardini, L.; Dahan-Moss, Y.; Koekemoer, L.L. 20-Hydroxyecdysone (20E) signaling as a promising target for the chemical control of malaria vectors. Parasites Vectors 2021, 14, 86. [Google Scholar] [CrossRef]
- Xu, K.-K.; Yang, W.-J.; Tian, Y.; Wu, Y.-B.; Wang, J.-J. Insulin signaling pathway in the oriental fruit fly: The role of insulin receptor substrate in ovarian development. General. Comp. Endocrinol. 2015, 216, 125–133. [Google Scholar] [CrossRef]
- Kannangara, J.R.; Mirth, C.K.; Warr, C.G. Regulation of ecdysone production in Drosophila by neuropeptides and peptide hormones. Open Biol. 2021, 11, 200373. [Google Scholar] [CrossRef]
- Dong, W.; Zhe, W.; XU, H.-q.; NIU, J.-z.; WANG, J.-j. Cloning and functional characterization of two peptidoglycan recognition protein isoforms (PGRP-LC) in Bactrocera dorsalis (Diptera: Tephritidae). J. Integr. Agric. 2020, 19, 3025–3034. [Google Scholar]
- Kampouraki, A.; Tsakireli, D.; Koidou, V.; Stavrakaki, M.; Kaili, S.; Livadaras, I.; Grigoraki, L.; Ioannidis, P.; Roditakis, E.; Vontas, J. Functional characterization of cytochrome P450s associated with pyrethroid resistance in the olive fruit fly Bactrocera oleae. Pestic. Biochem. Physiol. 2023, 191, 105374. [Google Scholar] [CrossRef]
- Bai, P.-P.; Chen, E.-H.; Shen, G.-M.; Wei, D.; Wei, D.-D.; Wang, J.-J. Inhibition of phenoloxidase activity delays development in Bactrocera dorsalis (Diptera: Tephritidae). Fla. Entomol. 2014, 97, 477–485. [Google Scholar] [CrossRef]
- Huang, Y.; Yu, Y.; Zhan, S.; Tomberlin, J.K.; Huang, D.; Cai, M.; Zheng, L.; Yu, Z.; Zhang, J. Dual oxidase Duox and Toll-like receptor 3 TLR3 in the Toll pathway suppress zoonotic pathogens through regulating the intestinal bacterial community homeostasis in Hermetia illucens L. PLoS ONE 2020, 15, e0225873. [Google Scholar] [CrossRef]
- Lima, L.F.; Torres, A.Q.; Jardim, R.; Mesquita, R.D.; Schama, R. Evolution of Toll, Spatzle and MyD88 in insects: The problem of the Diptera bias. BMC Genom. 2021, 22, 562. [Google Scholar] [CrossRef]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and applications of CRISPR-Cas9 for genome engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef]
- Bai, X.; Zeng, T.; Ni, X.Y.; Su, H.A.; Huang, J.; Ye, G.Y.; Lu, Y.Y.; Qi, Y.X. CRISPR/Cas9-mediated knockout of the eye pigmentation gene white leads to alterations in colour of head spots in the oriental fruit fly, Bactrocera dorsalis. Insect Mol. Biol. 2019, 28, 837–849. [Google Scholar] [CrossRef]
- Chen, H.; Sun, H.; Xie, J.; Yao, Z.; Zheng, W.; Li, Z.; Deng, Z.; Li, X.; Zhang, H. CRISPR/Cas9-induced Mutation of Sex Peptide Receptor Gene Bdspr Affects Ovary, Egg Laying, and Female Fecundity in Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). J. Insect Sci. 2023, 23, 2. [Google Scholar] [CrossRef]
- Pitt, A.R.; Kolch, W. 24 Chapter Genomics, Proteomics, and Metabolomics. In Medical Biochemistry-E-Book; Elsevier Health Sciences: Barcelona, Spain, 2022; Volume 333. [Google Scholar]
- Yang, Y.; Jiang, H.-B.; Liang, C.-H.; Ma, Y.-P.; Dou, W.; Wang, J.-J. Chromosome-level genome assembly reveals potential epigenetic mechanisms of the thermal tolerance in the oriental fruit fly, Bactrocera dorsalis. Int. J. Biol. Macromol. 2023, 225, 430–441. [Google Scholar] [CrossRef]
- Altstein, M.; Nässel, D.R. Neuropeptide Signaling in Insects. In Neuropeptide Systems as Targets for Parasite and Pest Control; Geary, T.G., Maule, A.G., Eds.; Springer: Boston, MA, USA, 2010; pp. 155–165. [Google Scholar] [CrossRef]
- Hou, Q.L.; Jiang, H.B.; Gui, S.H.; Chen, E.H.; Wei, D.D.; Li, H.M.; Wang, J.J.; Smagghe, G. A Role of Corazonin Receptor in Larval-Pupal Transition and Pupariation in the Oriental Fruit Fly Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Front. Physiol. 2017, 8, 77. [Google Scholar] [CrossRef]
- Gui, S.-H.; Jiang, H.-B.; Smagghe, G.; Wang, J.-J. The neuropeptides and protein hormones of the agricultural pest fruit fly Bactrocera dorsalis: What do we learn from the genome sequencing and tissue-specific transcriptomes? Peptides 2017, 98, 29–34. [Google Scholar] [CrossRef]
- Caplan, S.L.; Milton, S.L.; Dawson-Scully, K. A cGMP-dependent protein kinase (PKG) controls synaptic transmission tolerance to acute oxidative stress at the Drosophila larval neuromuscular junction. J. Neurophysiol. 2013, 109, 649–658. [Google Scholar] [CrossRef]
- Miyazaki, H.; Otake, J.; Mitsuno, H.; Ozaki, K.; Kanzaki, R.; Chieng, A.C.-T.; Hee, A.K.-W.; Nishida, R.; Ono, H. Functional characterization of olfactory receptors in the oriental fruit fly Bactrocera dorsalis that respond to plant volatiles. Insect Biochem. Mol. Biol. 2018, 101, 32–46. [Google Scholar] [CrossRef]
- Xu, L.; Jiang, H.-B.; Tang, K.-Y.; Yan, Y.; Schetelig, M.F.; Wang, J.-J. CRISPR-mediated mutagenesis of the odorant receptor co-receptor (Orco) gene disrupts olfaction-mediated behaviors in Bactrocera dorsalis. Insect Sci. 2022, 29, 1275–1286. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.H.; Zhang, Q.; Yang, R.L.; Yuan, G.R.; Wang, J.J.; Dou, W. RNAi-mediated knockdown of juvenile hormone acid O-methyltransferase disrupts larval development in the oriental fruit fly, Bactrocera dorsalis (Hendel). Pestic. Biochem. Physiol. 2022, 188, 105285. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhang, S.; Li, T.; Li, H.; Zhang, H.; Zheng, W. RNA sequencing identifies an ovary-enriched microRNA, miR-311-3p, involved in ovarian development and fecundity by targeting Endophilin B1 in Bactrocera dorsalis. Pest. Manag. Sci. 2023, 79, 688–700. [Google Scholar] [CrossRef]
- Wu, Z.; Yang, L.; He, Q.; Zhou, S. Regulatory Mechanisms of Vitellogenesis in Insects. Front. Cell Dev. Biol. 2020, 8, 593613. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, N. Chapter One—Ecdysteroid signalling in insects—From biosynthesis to gene expression regulation. In Advances in Insect Physiology; Adams, M.E., Ed.; Academic Press: Cambridge, MA, USA, 2021; Volume 60, pp. 1–36. [Google Scholar]
- Roh, G.-H.; Kendra, P.E.; Cha, D.H. Preferential attraction of oviposition-ready oriental fruit flies to host fruit odor over protein food odor. Insects 2021, 12, 909. [Google Scholar] [CrossRef]
- Kimbokota, F.; Hassanali, A.; Njagi, P.G.N. Potential attractants from three host plants of Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Int. J. Trop. Insect Sci. 2023, 43, 107–114. [Google Scholar] [CrossRef]
- Ren, L.; Zeng, L.; Lin, J.; Lu, Y.; Liu, C. Biological Control of Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Biol. Invasions Biol. Control Theory Pract. 2008, 413–440. [Google Scholar]
- Ren, L.; Ma, J.; Lv, Y.; Tong, Q.; Guo, H. Characterization of key off-odor compounds in thermal duck egg gels by GC-olfactometry-MS, odor activity values, and aroma recombination. LWT 2021, 143, 111182. [Google Scholar] [CrossRef]
- Xu, L.; Yu, X.; Li, M.; Chen, J.; Wang, X. Monitoring oxidative stability and changes in key volatile compounds in edible oils during ambient storage through HS-SPME/GC–MS. Int. J. Food Prop. 2017, 20, S2926–S2938. [Google Scholar] [CrossRef]
- Cornelius, M.L.; Duan, J.J.; Messing, R.H. Volatile host fruit odors as attractants for the oriental fruit fly (Diptera: Tephritidae). J. Econ. Entomol. 2000, 93, 93–100. [Google Scholar] [CrossRef]
- Zhu, X.; Li, Q.; Li, J.; Luo, J.; Chen, W.; Li, X. Comparative Study of Volatile Compounds in the Fruit of Two Banana Cultivars at Different Ripening Stages. Molecules 2018, 23, 2456. [Google Scholar] [CrossRef]
- Li, Y.-Y.; Liu, D.; Wen, P.; Chen, L. Detection of Volatile Organic Compounds by Antennal Lamellae of a Scarab Beetle. Front. Ecol. Evol. 2021, 9, 392–401. [Google Scholar] [CrossRef]
- Steinbrecht, R.A. Pore structures in insect olfactory sensilla: A review of data and concepts. Int. J. Insect Morphol. Embryol. 1997, 26, 229–245. [Google Scholar] [CrossRef]
- Francia, S.; Lodovichi, C. The role of the odorant receptors in the formation of the sensory map. BMC Biol. 2021, 19, 174. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Chen, Z.-S.; Zhang, D.-J.; Lu, Y.-Y. BdorOR88a Modulates the Responsiveness to Methyl Eugenol in Mature Males of Bactrocera dorsalis (Hendel). Front. Physiol. 2018, 9, 987. [Google Scholar] [CrossRef]
- Agnihotri, A.; Liu, N.; Xu, W. Chemosensory proteins (CSPs) in the cotton bollworm Helicoverpa armigera. Insects 2021, 13, 29. [Google Scholar] [CrossRef]
- Liu, Z.; Liang, X.-F.; Xu, L.; Keesey, I.W.; Lei, Z.-R.; Smagghe, G.; Wang, J.-J. An Antennae-Specific Odorant-Binding Protein Is Involved in Bactrocera dorsalis Olfaction. Front. Ecol. Evol. 2020, 8, 63. [Google Scholar] [CrossRef]
- Chen, J.; Li, S.-S.; Fang, S.-M.; Zhang, Z.; Yu, Q.-Y. Olfactory dysfunction and potential mechanisms caused by volatile organophosphate dichlorvos in the silkworm as a model animal. J. Hazard. Mater. 2022, 425, 127940. [Google Scholar] [CrossRef]
- Liu, Z.; Xie, Q.-P.; Guo, H.-W.; Xu, W.; Wang, J.-J. An odorant binding protein mediates Bactrocera dorsalis olfactory sensitivity to host plant volatiles and male attractant compounds. Int. J. Biol. Macromol. 2022, 219, 538–544. [Google Scholar] [CrossRef]
- Jang, S.; Kikuchi, Y. Impact of the insect gut microbiota on ecology, evolution, and industry. Curr. Opin. Insect Sci. 2020, 41, 33–39. [Google Scholar] [CrossRef]
- Ren, X.; Cao, S.; Akami, M.; Mansour, A.; Yang, Y.; Jiang, N.; Wang, H.; Zhang, G.; Qi, X.; Xu, P.; et al. Gut symbiotic bacteria are involved in nitrogen recycling in the tephritid fruit fly Bactrocera dorsalis. BMC Biol. 2022, 20, 201. [Google Scholar] [CrossRef] [PubMed]
- Jing, T.-Z.; Qi, F.-H.; Wang, Z.-Y. Most dominant roles of insect gut bacteria: Digestion, detoxification, or essential nutrient provision? Microbiome 2020, 8, 38. [Google Scholar] [CrossRef] [PubMed]
- Guentzel, M.N. Escherichia, Klebsiella, Enterobacter, Serratia, Citrobacter, and Proteus. In Medical Microbiology; Baron, S., Ed.; The University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- Wang, H.; Jin, L.; Zhang, H. Comparison of the diversity of the bacterial communities in the intestinal tract of adult Bactrocera dorsalis from three different populations. J. Appl. Microbiol. 2011, 110, 1390–1401. [Google Scholar] [CrossRef] [PubMed]
- Zeng, T.; Jaffar, S.; Xu, Y.; Qi, Y. The Intestinal Immune Defense System in Insects. Int. J. Mol. Sci. 2022, 23, 15132. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Wang, A.; Li, Y.; Cai, Z.; Lemaitre, B.; Zhang, H. The dual oxidase gene BdDuox regulates the intestinal bacterial community homeostasis of Bactrocera dorsalis. ISME J. 2016, 10, 1037–1050. [Google Scholar] [CrossRef] [PubMed]
- Raza, M.F.; Wang, Y.; Cai, Z.; Bai, S.; Yao, Z.; Awan, U.A.; Zhang, Z.; Zheng, W.; Zhang, H. Gut microbiota promotes host resistance to low-temperature stress by stimulating its arginine and proline metabolism pathway in adult Bactrocera dorsalis. PLoS Pathog. 2020, 16, e1008441. [Google Scholar] [CrossRef]
- Akami, M.; Ren, X.; Wang, Y.; Mansour, A.; Cao, S.; Qi, X.; Ngakou, A.; Ngane, R.A.N.; Niu, C.-Y. Host fruits shape the changes in the gut microbiota and development of Bactrocera dorsalis (Diptera: Tephritidae) larvae. Int. J. Trop. Insect Sci. 2022, 42, 2127–2141. [Google Scholar] [CrossRef]
- Zeng, T.; Bai, X.; Liu, Y.L.; Li, J.F.; Lu, Y.Y.; Qi, Y.X. Intestinal responses of the oriental fruit fly Bactrocera dorsalis (Hendel) after ingestion of an entomopathogenic bacterium strain. Pest. Manag. Sci. 2020, 76, 653–664. [Google Scholar] [CrossRef]
- Koul, B.; Chopra, M.; Lamba, S. Chapter 3—Microorganisms as biocontrol agents for sustainable agriculture. In Relationship Between Microbes and the Environment for Sustainable Ecosystem Services, Volume 1; Samuel, J., Kumar, A., Singh, J., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 45–68. [Google Scholar] [CrossRef]
- Shi, Z.; Wang, L.; Zhang, H. Low Diversity Bacterial Community and the Trapping Activity of Metabolites from Cultivable Bacteria Species in the Female Reproductive System of the Oriental Fruit Fly, Bactrocera dorsalis Hendel (Diptera: Tephritidae). Int. J. Mol. Sci. 2012, 13, 6266–6278. [Google Scholar] [CrossRef]
- Liu, S.H.; Chen, Y.; Li, W.; Tang, G.H.; Yang, Y.; Jiang, H.B.; Dou, W.; Wang, J.J. Diversity of Bacterial Communities in the Intestinal Tracts of Two Geographically Distant Populations of Bactrocera dorsalis (Diptera: Tephritidae). J. Econ. Entomol. 2018, 111, 2861–2868. [Google Scholar] [CrossRef]
- Noman, M.S.; Liu, L.; Bai, Z.; Li, Z. Tephritidae bacterial symbionts: Potentials for pest management. Bull. Entomol. Res. 2020, 110, 1–14. [Google Scholar] [CrossRef]
- Wang, M.; Cao, B.; Gao, Q.; Sun, Y.; Liu, P.; Feng, L.; Wang, L. Detection of Enterobacter sakazakii and other pathogens associated with infant formula powder by use of a DNA microarray. J. Clin. Microbiol. 2009, 47, 3178–3184. [Google Scholar] [CrossRef]
- Li, H.; Ren, L.; Xie, M.; Gao, Y.; He, M.; Hassan, B.; Lu, Y.; Cheng, D. Egg-surface bacteria are indirectly associated with oviposition aversion in Bactrocera dorsalis. Curr. Biol. 2020, 30, 4432–4440.e4434. [Google Scholar] [CrossRef]
- Taddei, A.; Reisenzein, H.; Mouttet, R.; Lethmayer, C.; Egartner, A.; Gottsberger, R.A.; Blümel, S.; Heiss, C.; Pohn, C.; Reynaud, P. Morphological and Molecular Identification Protocols for Bactrocera dorsalis: A Joint Validation Study. PhytoFrontiers™ 2023, 3, 186–198. [Google Scholar] [CrossRef]
- Choi, K.S.; Samayoa, A.C.; Hwang, S.-Y.; Huang, Y.-B.; Ahn, J.J. Thermal effect on the fecundity and longevity of Bactrocera dorsalis adults and their improved oviposition model. PLoS ONE 2020, 15, e0235910. [Google Scholar] [CrossRef] [PubMed]
- Fiaboe, K.K.; Kekeunou, S.; Nanga, S.N.; Kuate, A.F.; Tonnang, H.E.; Gnanvossou, D.; Hanna, R. Temperature-based phenology model to predict the development, survival, and reproduction of the oriental fruit fly Bactrocera dorsalis. J. Therm. Biol. 2021, 97, 102877. [Google Scholar]
- Jiang, F.; Liang, L.; Wang, J.; Zhu, S. Chromosome-level genome assembly of Bactrocera dorsalis reveals its adaptation and invasion mechanisms. Commun. Biol. 2022, 5, 25. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-X.; Chen, Y.; Lei, Q.; Peng, Y.-Y.; Jiang, H.-B. Sublethal Dose of β-Cypermethrin Impairs the Olfaction of Bactrocera dorsalis by Suppressing the Expression of Chemosensory Genes. Insects 2022, 13, 721. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Kang, F.; Zhan, G.; Ma, C.; Li, Y.; Wang, L.; Wei, Y.; Gao, X.; Li, Z.; Wang, Y. The Effects of a Cold Disinfestation on Bactrocera dorsalis Survival and Navel Orange Quality. Insects 2019, 10, 452. [Google Scholar] [CrossRef]
- Lin, K.W.; Lin, H.L.; Shiesh, C.C.; Hsu, Y.L.; Lin, C.H.; Chen, S.C.; Yeh, W.B. Cold treatment for guava fruits infested with oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae). Appl. Entomol. Zool. 2020, 55, 37–44. [Google Scholar] [CrossRef]
- Hsu, Y.-L.; Chen, S.-C.; Lin, K.-W.; Shiesh, C.-C.; Lin, C.-H.; Yeh, W.-B. Quarantine vapor heat treatment of papaya fruit for Bactrocera dorsalis and Bactrocera cucurbitae (Diptera: Tephritidae). J. Econ. Entomol. 2018, 111, 2101–2109. [Google Scholar] [CrossRef] [PubMed]
- Shelly, T. Effects of methyl eugenol and raspberry ketone/cue lure on the sexual behavior of Bactrocera species (Diptera: Tephritidae). Appl. Entomol. Zool. 2010, 45, 349–361. [Google Scholar] [CrossRef]
- Mandanayake, M.A.R.A.; Hee, A.K.W. Assessment of non-methyl eugenol-responding lines of Bactrocera dorsalis (Diptera: Tephritidae) males on lure response and mating. Bull. Entomol. Res. 2023, 113, 396–401. [Google Scholar] [CrossRef]
- Fan, Y.; Zhang, C.; Qin, Y.; Yin, X.; Dong, X.; Desneux, N.; Zhou, H. Monitoring the Methyl Eugenol Response and Non-Responsiveness Mechanisms in Oriental Fruit Fly Bactrocera dorsalis in China. Insects 2022, 13, 1004. [Google Scholar] [CrossRef]
- Mandanayake, M.A.R.A.; Shohaimi, S.; Ghani, N.I.A.; Hee, A.K.W. Establishment of non-methyl eugenol-responding lines from feral Oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera Tephritidae). Phytoparasitica 2023, 51, 425–436. [Google Scholar] [CrossRef]
- Raza, A.B.M.; Majeed, M.Z.; Ismail, M.; Qasim, Z. Field evaluation of IPM strategies against Bactrocera dorsalis PUJZ. Punjab Univ. J. Zool. 2023, 38, 59–64. [Google Scholar] [CrossRef]
- Tan, K.-H.; Wee, S.-L.; Nishida, R.; Shelly, T.E. Attraction of feral Bactrocera dorsalis males (Diptera: Tephritidae) to natural versus commercial sources of methyl eugenol. J. Asia-Pac. Entomol. 2021, 24, 1095–1100. [Google Scholar] [CrossRef]
- Bautista, R.C.; Mochizuki, N.; Spencer, J.P.; Harris, E.J.; Ichimura, D.M. Mass-Rearing of the Tephritid Fruit Fly Parasitoid Fopius arisanus (Hymenoptera: Braconidae)1. Biol. Control 1999, 15, 137–144. [Google Scholar] [CrossRef]
- Lahiri, S.; Orr, D. Chapter 11—Biological Control in Tomato Production Systems: Theory and Practice. In Sustainable Management of Arthropod Pests of Tomato; Wakil, W., Brust, G.E., Perring, T.M., Eds.; Academic Press: San Diego, CA, USA, 2018; pp. 253–267. [Google Scholar] [CrossRef]
- Nanga Nanga, S.; Hanna, R.; Gnanvossou, D.; Fotso Kuate, A.; Fiaboe, K.K.M.; Djieto-Lordon, C. Fruit Preference, Parasitism, and Offspring Fitness of Fopius arisanus (Hymenoptera: Braconidae) Exposed to Bactrocera dorsalis’ (Diptera: Tephritidae) Infested Fruit Species. Environ. Entomol. 2019, 48, 1286–1296. [Google Scholar] [CrossRef]
- Sime, K.R.; Daane, K.M.; Wang, X.G.; Johnson, M.W.; Messing, R.H. Evaluation of Fopius arisanus as a biological control agent for the olive fruit fly in California. Agric. For. Entomol. 2008, 10, 423–431. [Google Scholar] [CrossRef]
- Zheng, Y.; Song, Z.-W.; Zhang, Y.-P.; Li, D.-S. Ability of Spalangia endius (Hymenoptera: Pteromalidae) to Parasitize Bactrocera dorsalis (Diptera: Tephritidae) after Switching Hosts. Insects 2021, 12, 613. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.-D.; Ji, X.-C.; Han, Y.; Fu, B.-L.; Liu, K. Parasitism, Emergence, and Development of Spalangia endius (Hymenoptera: Pteromalidae) in Pupae of Different Ages of Bactrocera cucurbitae (Diptera: Tephritidae). J. Insect Sci. 2015, 15, 15. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Chen, J.; Niu, L.; Han, D.; Zhang, F.; Fu, Y. Field evaluation of Spalangia endius Walker in controlling Zeugodacus cucurbitae (Coquillett). Int. J. Pest. Manag. 2021, 67, 216–221. [Google Scholar] [CrossRef]
- van Lenteren, J.C. The state of commercial augmentative biological control: Plenty of natural enemies, but a frustrating lack of uptake. BioControl 2012, 57, 1–20. [Google Scholar] [CrossRef]
- Liang, J.; Tang, S.; Cheke, R.A.; Wu, J. Adaptive release of natural enemies in a pest-natural enemy system with pesticide resistance. Bull. Math. Biol. 2013, 75, 2167–2195. [Google Scholar] [CrossRef]
- Blissard, G.W.; Theilmann, D.A. Baculovirus Entry and Egress from Insect Cells. Annu. Rev. Virol. 2018, 5, 113–139. [Google Scholar] [CrossRef]
- Prasad, V.; Srivastava, S. Chapter 13—Insect Viruses. In Ecofriendly Pest Management for Food Security; Omkar, Ed.; Academic Press: San Diego, CA, USA, 2016; pp. 411–442. [Google Scholar] [CrossRef]
- Zhang, W.; Gu, Q.; Niu, J.; Wang, J.J. The RNA Virome and Its Dynamics in an Invasive Fruit Fly, Bactrocera dorsalis, Imply Interactions Between Host and Viruses. Microb. Ecol. 2020, 80, 423–434. [Google Scholar] [CrossRef]
- Iqbal, M.; Gogi, M.D.; Atta, B.; Nisar, M.J.; Arif, M.J.; Javed, N. Assessment of pathogenicity of Beauveria bassiana, Metarhizium anisopliae, Verticillium lecanii and Bacillus thuringiensis var. kurstaki against Bactrocera cucurbitae Coquillett (Diptera: Tephritidae) via diet-bioassay technique under controlled conditions. Int. J. Trop. Insect Sci. 2021, 41, 1129–1145. [Google Scholar] [CrossRef]
- Wang, D.; Liang, Q.; Chen, M.; Ye, H.; Liao, Y.; Yin, J.; Lü, L.; Lei, Y.; Cai, D.; Jaleel, W.; et al. Susceptibility of oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae) pupae to entomopathogenic fungi. Appl. Entomol. Zool. 2021, 56, 269–275. [Google Scholar] [CrossRef]
- Martínez-Barrera, O.Y.; Toledo, J.; Cancino, J.; Liedo, P.; Gómez, J.; Valle-Mora, J.; Montoya, P. Interaction Between Beauveria bassiana (Hypocreales: Cordycipitaceae) and Coptera haywardi (Hymenoptera: Diapriidae) for the Management of Anastrepha obliqua (Diptera: Tephritidae). J. Insect Sci. 2020, 20, 1–10. [Google Scholar] [CrossRef]
- Shishir, M.A.; Akter, A.; Bodiuzzaman, M.; Hossain, M.A.; Alam, M.M.; Khan, S.A.; Khan, S.N.; Hoq, M.M. Novel toxicity of Bacillus thuringiensis strains against the melon fruit fly, Bactrocera cucurbitae (Diptera: Tephritidae). Biocontrol Sci. 2015, 20, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Wakil, W.; Usman, M.; Piñero, J.C.; Wu, S.; Toews, M.D.; Shapiro-Ilan, D.I. Combined application of entomopathogenic nematodes and fungi against fruit flies, Bactrocera zonata and B. dorsalis (Diptera: Tephritidae): Laboratory cups to field study. Pest. Manag. Sci. 2022, 78, 2779–2791. [Google Scholar] [CrossRef]
- Heve, W.K.; Adjadeh, T.A.; Billah, M.K. Overview and future research needs for development of effective biocontrol strategies for management of Bactrocera dorsalis Hendel (Diptera: Tephritidae) in sub-Saharan Africa. Pest. Manag. Sci. 2021, 77, 4224–4237. [Google Scholar] [CrossRef]
- Godjo, A.; Chabi, N.; Zadji, L.; Dossou, P.; Batcho, O.; Baimey, H.; Bonou, W.; Sinzogan, A.A.C.; Bokonon-Ganta, A.; Decraemer, W.; et al. Evaluation of the ability of indigenous nematode isolates of Heterorhabditis taysearae and Steinernema kandii to control mango fruit fly Bactrocera dorsalis under laboratory, semi-field and field conditions in Northern Benin. Crop Prot. 2021, 149, 105754. [Google Scholar] [CrossRef]
- Agboka, K.M.; Tonnang, H.E.Z.; Abdel-Rahman, E.M.; Kimathi, E.; Mutanga, O.; Odindi, J.; Niassy, S.; Mohamed, S.A.; Ekesi, S. A systematic methodological approach to estimate the impacts of a classical biological control agent’s dispersal at landscape: Application to fruit fly Bactrocera dorsalis and its endoparasitoid Fopius arisanus. Biol. Control 2022, 175, 105053. [Google Scholar] [CrossRef]
- Ndlela, S.; Mohamed, S.A.; Azrag, A.G.A.; Ndegwa, P.N.; Ong’amo, G.O.; Ekesi, S. Interactions between Two Parasitoids of Tephritidae: Diachasmimorpha longicaudata (Ashmead) and Psyttalia cosyrae (Wilkinson) (Hymenoptera: Braconidae), under Laboratory Conditions. Insects 2020, 11, 671. [Google Scholar] [CrossRef] [PubMed]
- Labaude, S.; Griffin, C.T. Transmission Success of Entomopathogenic Nematodes Used in Pest Control. Insects 2018, 9, 72. [Google Scholar] [CrossRef] [PubMed]
- Toledo, J.; Morán-Aceves, B.M.; Ibarra, J.E.; Liedo, P. Can Entomopathogenic Nematodes and Their Symbiotic Bacteria Suppress Fruit Fly Pests? A Review. Microorganisms 2023, 11, 1682. [Google Scholar] [CrossRef]
- Diouf, E.G.; Brévault, T.; Ndiaye, S.; Faye, E.; Chailleux, A.; Diatta, P.; Piou, C. An agent-based model to simulate the boosted Sterile Insect Technique for fruit fly management. Ecol. Model. 2022, 468, 109951. [Google Scholar] [CrossRef]
- Pérez-Staples, D.; Díaz-Fleischer, F.; Montoya, P. The Sterile Insect Technique: Success and Perspectives in the Neotropics. Neotrop. Entomol. 2021, 50, 172–185. [Google Scholar] [CrossRef]
- Cladera, J.L.; Vilardi, J.C.; Juri, M.; Paulin, L.E.; Giardini, M.C.; Gómez Cendra, P.V.; Segura, D.F.; Lanzavecchia, S.B. Genetics and biology of Anastrepha fraterculus: Research supporting the use of the sterile insect technique (SIT) to control this pest in Argentina. BMC Genet. 2014, 15 (Suppl. S2), S12. [Google Scholar] [CrossRef] [PubMed]
- Mau, R.; Jang, E.; Vargas, R. The Hawaii area-wide fruit fly pest management programme. In Area-Wide Control of Insect Pests; Springer: Berlin/Heidelberg, Germany, 2007; pp. 671–683. [Google Scholar]
- Haq, I.U.; Abd-Alla, A.; Tomas, U.S.; Meza, J.S.; Bourtzis, K.; Cáceres, C. Cryopreservation of the Mediterranean fruit fly (Diptera: Tephritidae) VIENNA 8 genetic sexing strain: No effect on large scale production of high quality sterile males for SIT applications. PLoS ONE 2019, 14, e0211259. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Santos, E.; Rendon, P.; Gouvi, G.; Zacharopoulou, A.; Bourtzis, K.; Cáceres, C.; Bloem, K. A Novel Genetic Sexing Strain of Anastrepha ludens for Cost-Effective Sterile Insect Technique Applications: Improved Genetic Stability and Rearing Efficiency. Insects 2021, 12, 499. [Google Scholar] [CrossRef]
- Sollazzo, G.; Gouvi, G.; Nikolouli, K.; Cancio Martinez, E.I.; Schetelig, M.F.; Bourtzis, K. Temperature Sensitivity of Wild-Type, Mutant and Genetic Sexing Strains of Ceratitis capitata. Insects 2022, 13, 943. [Google Scholar] [CrossRef]
- Robinson, A.S. Genetic sexing strains in medfly, Ceratitis capitata, sterile insect technique programmes. Genetica 2002, 116, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Aluja, M.; Norrbom, A. Fruit Flies (Tephritidae): Phylogeny and Evolution of Behavior; CRC Press: Boca Raton, FL, USA, 1999. [Google Scholar]
- Hendrichs, J.; Robinson, A.; Cayol, J.; Enkerlin, W. Medfly areawide sterile insect technique programmes for prevention, suppression or eradication: The importance of mating behavior studies. Fla. Entomol. 2002, 85, 1–13. [Google Scholar] [CrossRef]
- Kaneko, Y.; Szallasi, A. Transient receptor potential (TRP) channels: A clinical perspective. Br. J. Pharmacol. 2014, 171, 2474–2507. [Google Scholar] [CrossRef]
- Su, H.; Bai, X.; Zeng, T.; Lu, Y.; Qi, Y. Identification, characterization and expression analysis of transient receptor potential channel genes in the oriental fruit fly, Bactrocera dorsalis. BMC Genom. 2018, 19, 674. [Google Scholar] [CrossRef]
- Nesterov, A.; Spalthoff, C.; Kandasamy, R.; Katana, R.; Rankl, N.B.; Andrés, M.; Jähde, P.; Dorsch, J.A.; Stam, L.F.; Braun, F.-J.; et al. TRP Channels in Insect Stretch Receptors as Insecticide Targets. Neuron 2015, 86, 665–671. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.-J.; Guo, D.; Wang, L.-X.; Niu, C.-D.; Wu, S.-F.; Zhang, Y.V.; Gao, C.-F. Function of Transient Receptor Potential-Like Channel in Insect Egg Laying. Front. Mol. Neurosci. 2022, 15, 823563. [Google Scholar] [CrossRef]
- Wang, R.; Gao, B.; Zhang, Q.; Qu, C.; Luo, C. Knockdown of TRPV gene Nanchung decreases resistance to the novel pyropene insecticide, afidopyropen, in Bemisia tabaci. Int. J. Biol. Macromol. 2023, 224, 1566–1575. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.F.; Niu, J.Z.; Jiang, X.Z.; Yang, W.J.; Shen, G.M.; Wei, D.; Smagghe, G.; Wang, J.J. Influence of various stressors on the expression of core genes of the small interfering RNA pathway in the oriental fruit fly, Bactrocera dorsalis. Insect Sci. 2017, 24, 418–430. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, M.; Zhang, H. RNA interference of four genes in adult Bactrocera dorsalis by feeding their dsRNAs. PLoS ONE 2011, 6, e17788. [Google Scholar] [CrossRef]
- Dong, Y.-C.; Wang, Z.-J.; Chen, Z.-Z.; Clarke, A.R.; Niu, C.-Y. Bactrocera dorsalis male sterilization by targeted RNA interference of spermatogenesis: Empowering sterile insect technique programs. Sci. Rep. 2016, 6, 35750. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Fu, W.-B.; Si, F.-L.; Yan, Z.-T.; Zhang, Y.-J.; He, Q.-Y.; Chen, B. UDP-glycosyltransferase genes and their association and mutations associated with pyrethroid resistance in Anopheles sinensis (Diptera: Culicidae). Malar. J. 2019, 18, 62. [Google Scholar] [CrossRef]
- Chen, M.-L.; Zhang, S.-X.; Guo, P.-Y.; Qin, Q.-S.; Meng, L.-W.; Yuan, G.-R.; Wang, J.-J. Identification and characterization of UDP-glycosyltransferase genes and the potential role in response to insecticides exposure in Bactrocera dorsalis. Pest. Manag. Sci. 2023, 79, 666–677. [Google Scholar] [CrossRef]
- Moon, T.T.; Maliha, I.J.; Khan, A.A.M.; Chakraborty, M.; Uddin, M.S.; Amin, M.R.; Islam, T. CRISPR-Cas Genome Editing for Insect Pest Stress Management in Crop Plants. Stresses 2022, 2, 493–514. [Google Scholar] [CrossRef]
- Wang, X.-F.; Chen, Z.; Wang, X.-B.; Xu, J.; Chen, P.; Ye, H. Bacterial-mediated RNAi and functional analysis of Natalisin in a moth. Sci. Rep. 2021, 11, 4662. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Bartalska, K.; Audsley, N.; Yamanaka, N.; Yapici, N.; Lee, J.-Y.; Kim, Y.-C.; Markovic, M.; Isaac, E.; Tanaka, Y.; et al. MIPs are ancestral ligands for the sex peptide receptor. Proc. Natl. Acad. Sci. USA 2010, 107, 6520–6525. [Google Scholar] [CrossRef]
- Zheng, W.; Liu, Y.; Zheng, W.; Xiao, Y.; Zhang, H. Influence of the silencing sex-peptide receptor on Bactrocera dorsalis adults and offspring by feeding with ds-spr. J. Asia-Pac. Entomol. 2015, 18, 477–481. [Google Scholar] [CrossRef]
Figure 1.
The life cycle of B. dorsalis.

Figure 2.
Distribution of B. dorsalis in China. The map information was produced using the geospatial data cloud https://www.gscloud.cn (accessed on 19 August 2023) and datav.aliyun.com (accessed on 19 August 2023). The spatial analysis was accessed through ArcGIS version (10.7) and Mapshaper, which were applied on 19 August.
Figure 2.
Distribution of B. dorsalis in China. The map information was produced using the geospatial data cloud https://www.gscloud.cn (accessed on 19 August 2023) and datav.aliyun.com (accessed on 19 August 2023). The spatial analysis was accessed through ArcGIS version (10.7) and Mapshaper, which were applied on 19 August.

Figure 3.
Key factors involved in the invasion biology of B. dorsalis.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jaffar, S.; Rizvi, S.A.H.; Lu, Y. Understanding the Invasion, Ecological Adaptations, and Management Strategies of Bactrocera dorsalis in China: A Review. Horticulturae 2023, 9, 1004. https://doi.org/10.3390/horticulturae9091004
AMA Style
Jaffar S, Rizvi SAH, Lu Y. Understanding the Invasion, Ecological Adaptations, and Management Strategies of Bactrocera dorsalis in China: A Review. Horticulturae. 2023; 9(9):1004. https://doi.org/10.3390/horticulturae9091004
Chicago/Turabian StyleJaffar, Saleem, Syed Arif Hussain Rizvi, and Yongyue Lu. 2023. "Understanding the Invasion, Ecological Adaptations, and Management Strategies of Bactrocera dorsalis in China: A Review" Horticulturae 9, no. 9: 1004. https://doi.org/10.3390/horticulturae9091004
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.