
Cistus albidus L.—Review of a Traditional Mediterranean Medicinal Plant with Pharmacological Potential



1
Department of Engineering, Area of Agroforestry, Miguel Hernández University, 03312 Orihuela, Spain
2
Institute for Research, Development and Innovation in Health Biotechnology, Miguel Hernández University, 03202 Elche, Spain
3
Department of Pharmacy, Elche University Hospital-FISABIO, 03203 Elche, Spain
*
Author to whom correspondence should be addressed.
Plants 2023, 12(16), 2988; https://doi.org/10.3390/plants12162988
Submission received: 12 July 2023
/
Revised: 11 August 2023
/
Accepted: 11 August 2023
/
Published: 18 August 2023
(This article belongs to the Special Issue Bioactive Compounds in Plants)
Abstract
:Cistus albidus L. (Cistaceae) is a medicinal plant that has been used therapeutically since ancient times in the Mediterranean basin for its important pharmacological properties. The ability of C. albidus to produce large quantities of a wide range of natural metabolites makes it an attractive source of raw material. The main constituents with bioactive functions that exert pharmacological effects are terpenes and polyphenols, with more than 200 identified compounds. The purpose of this review is to offer a detailed account of the botanical, ethnological, phytochemical, and pharmacological characteristics of C. albidus with the aim of encouraging additional pharmaceutical investigations into the potential therapeutic benefits of this medicinal plant. This review was carried out using organized searches of the available literature up to July 2023. A detailed analysis of C. albidus confirms its traditional use as a medicinal plant. The outcome of several studies suggests a deeper involvement of certain polyphenols and terpenes in multiple mechanisms such as inflammation and pain, with a potential application focus on neurodegenerative diseases and disorders. Other diseases such as prostate cancer and leukemia have already been researched with promising results for this plant, for which no intoxication has been reported in humans.
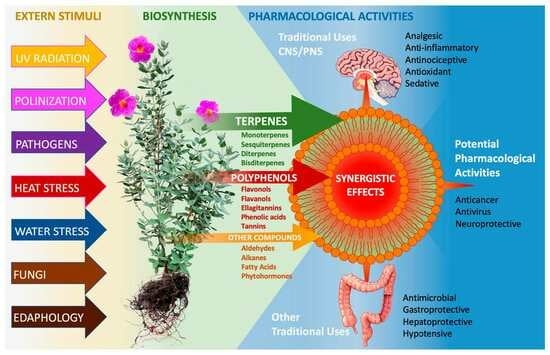
1. Introduction
Cistus albidus is one of the approximately 20 species of the Cistus genus [1]. The genus’ name is derived from the ancient Greek term kistos [2]. It is supposed that the name alludes to the woody capsule fruits. Evergreen in its Mediterranean homeland and between 50 and 250 centimeters tall [3], this shrub is called albidus, not because of the colour of its flowers, but because its leaves are finely covered with white hair (trichomes) [4]. For its optimal development, it needs calcareous, sandy, or siliceous soils, as found in the garrigue of the Mediterranean [5,6]. Here, it can grow in large groups (known as jarales in Spanish) (Figure 1) and sometimes invade adjoining areas. It does not make high demands on the soil in terms of nutrients [6], but it needs permeability as it does not tolerate stagnant soils.
The characteristic feature of C. albidus is that its leaves are covered on both sides with dense hairiness made up of a combination of glandular and non-glandular trichomes (Figure 2). The glandular trichomes can be stellate or solitary; they are elongated and produce and secrete a resin rich in metabolites of pharmacological interest [7], such as flavonoid aglycones, glycosides, and terpenoids, including the characteristic labdane-type diterpenes described in Table 1 and Table 2. It is because of these compounds that this species was traditionally used in popular medicine according to its pharmacological action as an anti-inflammatory, antimicrobial, antinociceptive, and sedative [8,9,10].
Currently, its use as a medicinal plant is vestigial; however, its phytochemical composition, especially the combination of some terpenoids and polyphenols, makes it a promising species with many pharmacological activities. Pharmacological studies on extracts of C. albidus have shown their antioxidant, antibacterial, antifungal [5,11,12], antinociceptive [13], and anticarcinogenic [5,14] activity, in addition to other potential activities discussed later in this review.
Nowadays, however, C. albidus is known more for its ability to withstand severe summer stress situations to repopulate degenerate areas [15,16,17,18] than for its important pharmacological properties. Due to its ability to produce large amounts of secondary metabolites, it becomes an attractive model for elucidation of the biosynthetic pathways involved in resisting climatic adversities [19,20,21,22]. The pathways that lead to the production of terpenes and polyphenols have been investigated on many occasions, and the substances have been analyzed in isolation. However, in the author´s opinion, the potential of C. albidus does not lie in the independent substances that compose it, but in the interaction among them.
2. Distribution
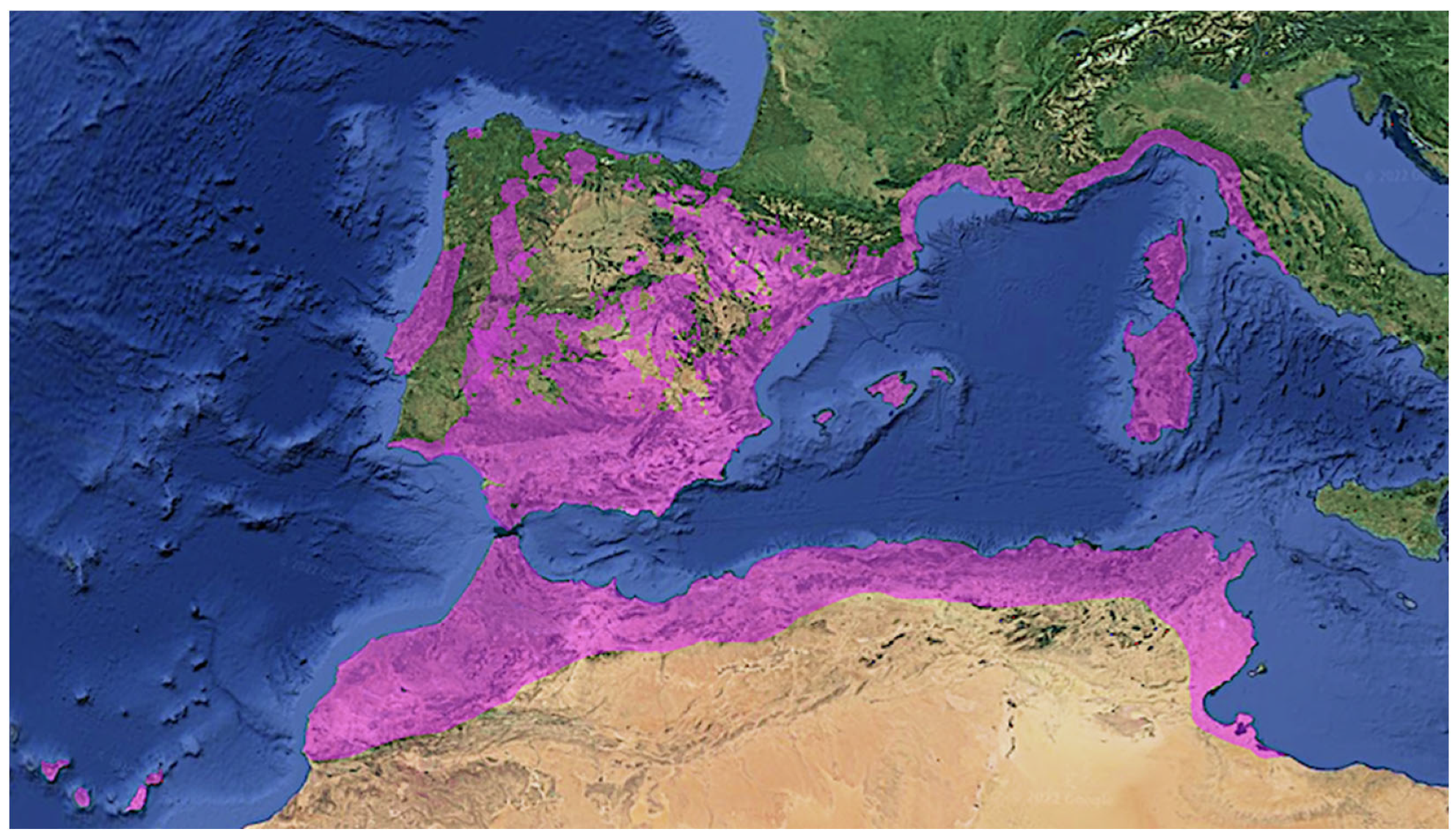
Cistus is a characteristic genus of the Mediterranean flora. The rockrose family has spread throughout the western Mediterranean. The genus is now believed to comprise approximately 20 species, of which 16 are native to Europe [1]. Some species of Cistus are endemic, while others are widespread in the Iberian Peninsula, the Canary Islands, Northwest Africa, Italy, Greece, France, or Turkey [1,21].
The often-coexisting species C. albidus L., C. crispus L., C. creticus L., and C. heterophyllus Desf. are native only to the western part of the Mediterranean, with the exception of C. criticus, which also reaches the eastern Mediterranean and the Black Sea coasts and the Crimean peninsula [18]. As can be seen in Figure 3, the currently known distribution of C. albidus includes the Iberian Peninsula, the Canary Islands, and the western Mediterranean. The northernmost population, by far, of C. albidus is found near Lake Garda in Italy [23]. It grows from sea level to 1300 m [24] and tolerates temperatures down to −12 °C, having an even lower thermal threshold [23].
3. Systematics
Cistus is one of eight genera found within the rockrose family (Cistaceae), which is a part of the Malvales order. The Cistaceae encompasses approximately 180 species [25]. Within this family, genera that are native to the Mediterranean basin include Cistus, Halimium, Fumana, Tuberaria, and Helianthemum, while Crocanthemum, Hudsonia, and Lechea are endemic to the American continent [26]. The Cistus genus is further categorized into the subgenus Cistus, which includes nine species. In the Iberian Peninsula, this subgenus contains species such as C. albidus L., C. crispus L., C. creticus L., and C. heterophyllus Desf., which often coexist. Among the species C. crispus, C. creticus, and C. heterophyllus, C. albidus represents one of the four paraphyletic species within the subgenus Cistus [18] (Figure 4). DNA sequence and pollen analysis showed a close evolutionary relationship of C. creticus and C. albidus, with C. crispus and C. heterophyllus more distantly related taxa [1,11,21]. Calculations have determined that the species C. albidus, C. creticus, and C. heterophyllus separated 0.19 Ma ago [21].
All representatives of this genus have a similar chromosome number (2n = 18), which has facilitated hybridization between species [11]. Nowadays, more than twenty natural interspecific hybrids are known, in addition to numerous cultivars of hybrid origin used for ornamental purpose [1,18]. As a consequence of this scenario, although the main characteristics of C. albidus are easily identifiable, they can be confused with these species due to their similarity of characteristics, especially the inflorescences.
4. Botanical Characteristics
From the morphological point of view, C. albidus is characterized by having five sepals, five pink to purple petals, up to 150 yellow stamens, pollen with a thin exine up to 1.4 μm thick, a long style that reaches or exceeds the stamens, and three cellular polysperm capsules. The hermaphroditic, actinomorphic, and hypogynous flowers, which appear from February to July, normally reach a diameter of four to six centimeters and develop individually or in umbels of usually five to seven [25]. The five sepals are ovate-lanceolate and hairy. The five petals, on the other hand, are delicate and slightly wrinkled. The flowers open in the morning and after a few hours the plant loses its petals. The flowers rarely last more than a day. A single adult bush can produce more than 1000 flowers per flowering period, depending on age [27].
The ovate-lanceolate leaves are arranged opposite each other and are usually 20–50 mm long and 8–30 mm wide; however, specimens with leaves exceeding 100 mm long and more than 50 mm wide have also been found by the authors. They are sessile, with a smooth margin, but may occasionally have a slightly wavy edge. Foliar veins are composed of three to five principal veins with a strong central vein. Adaxis veins are sunken, while veins on the abaxial surface are raised (Figure 2).
Due to its morphological adaptation to an extremely dry climate within Mediterranean regions and a pronounced resistance to abiotic stress in general, C. albidus could be considered a malacophilous xerophyte. During dry periods, these plants reduce the growth of relatively long and wide hairy leaves until only short, narrow leaves remain. At the same time, the hairiness (trichomes) of the leaves of these plants is increased. In general, these processes significantly reduce respiration, which is an effective protection mechanism against dehydration [28,29].
Vegetative Development
C. albidus has developed mechanisms that allow it to withstand severe summer droughts. These mechanisms consist in part of a reduction in leaf area and angle [28], combined with an increase in root mass per leaf area and modification of stomatal conductance [30]. On the other hand, the link between tocopherols and jasmonates appears to be primarily responsible for the regulation of biotic and abiotic stress responses [20]. It was observed that, under water stress, C. albidus increases enzymes related to redox homeostasis, such as oxidized ascorbate reductase, glyoxalase, superoxide dismutase, and isoflavone reductase, which was related to a reduction in oxidative stress in C. albidus exposed to drought and the ability to recover quickly after re-irrigation [19]. The interaction of these mechanisms allows this species to resist the adverse Mediterranean climate conditions with only little photoinhibition [20].
Furthermore, C. albidus belongs to the nanophanerophytes (a subgroup of phanerophytes). These are plants whose overwintering buds are above the level of the snow cover. In contrast to the macrophanerophytes, they do not rise above the level of the surrounding vegetation. They are therefore partially sheltered from the wind. Due to this fact, a more than 2 m high specimen was discovered in the Valle y Carrascoy Regional Park in Murcia, surrounded by shrubs up to 3 m high.
The vegetative development of this species, which normally lives for about 14 years [31] and was found to reach 17 [32] or even 25 years [27], is characterized by two types of lateral shoots, dolichoblasts and brachyblasts. Dolichoblasts are long shoots with large leaves, which are produced when climatic conditions are benign (availability of water and absence of frost), which is usually between the end of February to May and from September to December. Brachyblasts are short shoots that develop throughout the year in the axils of leaves of dolichoblast shoots [33]. The fall of the leaves of this marcescent species is acropetic.
Sexual reproduction begins at about the age of one year. The flowers of this partially self-incompatible species [34] normally last around 12 h but may last up to two days on the plant, especially in rainy weather with high relative air humidity. This seems to be due to the fact that the apoidea, its main pollinators [34], do not fly in humid environmental conditions. It has been further shown that zeatin is the substance that modulates the speed of floral development, depending on the age of the plants [22].
Fruiting takes place from May to August. The capsules contain an average of about 80 seeds, with exceptions found by the authors from fewer than 10 seeds to more than 140, and generally mature from August to December [33,34], but in warmer regions such as the Spanish Levant, for example, the first capsules usually mature at the end of May. Mature capsules spread their seeds close to parent plants as they lack expansion mechanisms. Studies suggest that C. albidus seeds experience a combination of physical and physiological dormancy [35]. Although physical dormancy was broken and water was available, the seeds seemed able to partially control their dormancy and germination capacity [35]. Under optimal conditions, germination takes between five and ten days and is epigeal [36].
Like other plants typical of fire-prone regions, C. albidus is generally considered pyrophytic [37], especially since the heat generated by fire is thought to facilitate breaking physical dormancy due to the hard seed coat [31,38,39]; thus, it is one of the first shrubs to emerge after a fire. Development is therefore rapid within the first five years and then progressively slows down [31]. However, the scarification of the seeds, by soil particles, (through dragging by water flow) also softens the hard cover of the seeds, making them permeable to water [40], thus overcoming physical dormancy [40,41]. In addition, it is common to find this species along the edges of watercourses with temporary flows. This seems to be its dispersal strategy since forest fires do not facilitate the spatial expansion of this species. Very specific conditions must be met for the seeds to viably survive forest fires and also break the integument. For this reason, C. albidus could be considered an opportunistic pioneer plant.
Regarding the influence of the soil on the development of C. albidus, together with C. creticus, it is the only taxon of the purple-pink clade capable of growing independently in calcareous and acidic soils. However, C. albidus grows best on calcareous soils in Mediterranean climates [5,42]. Studies reported that no significant qualitative or quantitative differences were found in the polyphenolic profile between the cultivation of C. albidus in different types of soil [5,6], while the concentration of terpenes was influenced by soil conditions, showing lower yields in calcareous soils [43]. However, higher concentrations of polyphenols were not associated with lower soil fertility [44]. This suggests that the genetic influence of this species on the biosynthesis of phytochemicals may be stronger than the influence of soil parameters. This was confirmed by a recent study, where C. albidus exhibited a low translocation of Pb and Cd to aerial parts from heavy-metal-contaminated soil [17], making this species also suitable for plantations under problematic soil conditions.
5. Phytochemical Constituents
The main constituents of C. albidus-derived products belong to the groups of terpenes and polyphenols. Other organic compounds have also been detected. To date, more than 200 secondary metabolites have been reported in C. albidus samples. In this review, 153 terpenoids, including 31 monoterpenes, 109 sesquiterpenes, 9 diterpenes, and 3 tetraterpenes, and their respective derivatives were found. In addition, 58 polyphenols, including 19 phenolic acids, 17 flavonols, 11 flavanols, 3 ellagitannins, 3 anthocyanins, 2 flavones, 1 anthocyanidin, 1 flavanone, and 1 hydrolysable tannin, were found. Moreover, 8 fatty acids, 7 alkanes, and various other compounds were found in the literature and discussed in the present study.
The irregular presence of reported compounds–analyses were often very heterogeneous—is possibly due to seasonal variation and the analytical methods used. Secondary metabolites reported so far have been determined under a wide variety of conditions, making it sometimes difficult to compare among the studies. For example, some metabolites, such as punicalagin derivatives or some diterpenes, were only found in a small number of studies [5,11]. The results depend on multiple factors, such as the method of analysis, the season and hours of the day of collection, and the type of processing, among others, and some of these factors are not detailed in many studies. However, on the other hand, there are a number of compounds that practically all authors found in significant amounts, being thus characteristic of this species. These more common compounds will be highlighted in the following sections.
The various compound structures detected in C. albidus are described below, headed by the terpenes and paying particular attention to the essential oils (mono- and sesquiterpenes). Phenylpropanoids detected in essential oils are also briefly discussed. Furthermore, di- and tetraterpenes are described. The second large group of substances, polyphenols, are examined more closely, especially flavonols, flavanols, ellagitannins, and phenolic acids. The carbonyl compounds and alkanes found so far are then listed. Finally, proven phytohormones and various fatty acids are described. However, the latter is not part of the secondary metabolism but is included here for its supposed importance as a potential bioavailability enhancer.
5.1. Terpenes
5.1.1. Mono- and Sesquiterpenes from the Essential Oils
Terpenes, a very large but heterogeneous group of naturally occurring secondary metabolites in C. albidus, represent the group with the most compounds identified in this species. Among them, the most abundant were the sesquiterpenes; they were, in addition, the largest class of identified compounds in C. albidus. While monoterpenes were found to contribute, to a certain extent, together with aldehydes, to the characteristic odor of this species (they are among the main components of floral aromas), the sesquiterpenes play a signaling role in the defense mechanisms of this species, acting as herbivore repellents or through the attraction of predators. Diterpenes are also synthesized for defense purposes and serve as precursors for vitamins and hormones such as tocopherols and gibberellins. Finally, tetraterpenes contribute to the pigmentation of flowers and fruits, playing an essential role in the pollination and distribution of seeds. As an antioxidant, it protects C. albidus from oxidative stress caused by adverse growing conditions [30].
The ISO definition for essential oils is “Product obtained from a natural raw material of plant origin, by steam distillation, by mechanical processes from the epicarp of citrus fruits, or by dry distillation, after separation of the aqueous phase—if any—by physical processes” [45].
Essential oils from fresh aerial parts of C. albidus were generally obtained in low yields of 0.01–0.1% (w/w) [46,47,48] by steam distillation. The seeds, in particular, contain very small amounts of essential oil, sometimes insufficient to be analyzed, with a yield of less than 0.01% [48]. The concentration of terpenes depends fundamentally on soil conditions (the more calcareous, the lower the yield), climatic factors, and the season [43]. Analysis of seasonal variation in terpene composition shows strong interannual variability, with the highest emission rates in autumn and spring and the lowest in summer and winter, leading to maximum values of stored terpenes in autumn and winter, while the spring and summer values showed minimum levels [47]. Table 1 shows the list of the terpenoids identified in C. albidus samples, including information about their previously published pharmacological activity. The compounds most frequently found in the analyses of the extracted terpenes (w/w), and therefore representative of this species, whether in leaves, pollen, flowers, flowering tops, or stems, are α-zingiberene (7.4–20.7%), aromadendrene (1.0–10.6%), ar-curcumene (8.3–13.2%), and germacrene D (1.0–7.9%) [46,48,49,50,51,52,53]. Among the terpenes in leaves, flowering tops, and flowers, monoterpenes were only present in small quantities and sometimes only in traces, while sesquiterpenes were the most abundant [46,48,49].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Identified terpenes in Cistus albidus. Table includes the structure for each compound along with the references in which it was identified and its pharmacological activity (including the references for this activity). 🌱: aerial parts, including leaves and twigs; ✿: flowering tops, flowers, petals, and sepals; 𐩕: pollen. n/a: reliable data are not available.
Table 1.
Identified terpenes in Cistus albidus. Table includes the structure for each compound along with the references in which it was identified and its pharmacological activity (including the references for this activity). 🌱: aerial parts, including leaves and twigs; ✿: flowering tops, flowers, petals, and sepals; 𐩕: pollen. n/a: reliable data are not available.
| No | Compound | Structure/Class | Presence in | Analytical Reference | Pharmacology |
|---|---|---|---|---|---|
| 1 | cis-α-Bergamotene | polycyclic monoterpene hydrocarbon | 🌱✿ | [48,49,53] | n/a |
| 2 | trans-α-Bergamotene | 🌱 | [47,48] | ||
| 3 | Borneol | polycyclic monoterpene alcohol | 🌱 | [52] | blood brain barrier (BBB) permeability improvement, intercellular tight junction (TJ) loosening [54] |
| 4 | Camphor | oxygenated polycyclic monoterpene | 🌱 | [50] | analgesic, antinociceptive [55]; antimicrobial, antiviral [56]; anticancer [57]; antitussive [58]; skin penetration enhancer [59]; |
| 5 | Carene | polycyclic monoterpene hydrocarbon | 🌱✿ | [52] | antiviral [60]; enhances bone mineralization [61]; anti-inflammatory [62]; |
| 6 | Carvacrol | monocyclic monoterpene alcohol | 🌱✿ | [49] | antibacterial (64); antifungal [63]; antioxidant [64]; anticancer [65]; anti-inflammatory, analgesic [66]; antiobesity [67]; hepatoprotective [68]; spasmolytic [69]; vasorelaxant [70]; |
| 7 | β-Cyclocitral | oxygenated monocyclic monoterpene | 🌱 | [53] | n/a |
| 8 | p-Cymene | monocyclic monoterpene hydrocarbon | 🌱 | [49,50] | anti-inflamatory, antinociceptive, antioxidant [71]; antidiabetic [72]; |
| 9 | p-Cymenene | monocyclic monoterpene hydrocarbon | 🌱✿ | [50] | n/a |
| 10 | Isobornyl formate | oxygenated polycyclic monoterpene | 🌱 | [50] | n/a |
| 11 | (D-)Limonene | monocyclic monoterpene hydrocarbon | 🌱✿ | [47,48,49,50] | anticancer, anticholesterol [73]; antidepressant [74]; |
| 12 | Linalool | acyclic monoterpene alcohol | 🌱✿ | [51,52] | antibacterial, antifungal [75]; anxiolytic [76]; anticancer, antioxidant [77]; analgesic [78]; anti-inflammatory [79]; |
| 13 | cis-linalool oxide | oxygenated heteromonocyclic monoterpene | [51] | n/a | |
| 14 | Myrcene | acyclic monoterpene hydrocarbon | 🌱 | [49,51] | analgesic, antinociceptive [80]; |
| 15 | Neryl acetate | acyclic monoterpene hydrocarbon | 🌱✿ | [53] | n/a |
| 16 | (E)-Ocimene | acyclic monoterpene hydrocarbon | 🌱✿ | [49] | anticancer [81]; anticonvulsant [82]; |
| 17 | (Z)-β-Ocimene | [49] | anticancer [81]; antibacterial [83]; | ||
| 18 | α-Phellandrene | monocyclic monoterpene hydrocarbon | 🌱 | [49] | antifungal [84]; antidepressant [85]; anti-inflammatory, antihyperalgesic [85,86], analgesic, antinociceptive [86]; anticancer [87]; |
| 19 | β-Phellandrene | ✿ | [49] | n/a | |
| 20 | α-Pinene | polycyclic monoterpene hydrocarbon | 🌱 | [47,49,51] | antifungal, anti-inflammatory, antioxidant [88]; anticancer [89]; anti-Leishmania [90]; gastroprotective [91,92]; antibacterial [93]; antiviral [94]; neuroprotective [95]; |
| 21 | β-Pinene | 🌱✿ | [47] | anticancer [96]; antimicrobial [97]; gastroprotective [92]; neuroprotective [98]; | |
| 22 | Piperitone | oxygenated monocyclic monoterpene | 𐩕 | [49] | n/a |
| 23 | Sabinene | polycyclic monoterpene hydrocarbon | 🌱 | [49] | n/a |
| 24 | cis-Sabinene hydrate | oxygenated polycyclic monoterpene | [49] | ||
| 25 | Safranal | oxygenated monocyclic monoterpene | 🌱 | [53] | antioxidant [99]; antimicrobial [100]; anticonvulsant [101]; antidepressant, anxiolytic [102]; gastroprotective [103]; |
| 26 | α-Terpinene | monocyclic monoterpene hydrocarbon | 🌱 | [49] | antioxidant [104]; antimicrobial [105]; |
| 27 | Δ-Terpinene | 🌱✿ | [52] | n/a | |
| 28 | γ-Terpinene | ✿ | [49] | antimicrobial [105]; | |
| 29 | α-Terpineol | monocyclic monoterpene alcohol | ✿𐩕 | [49,50,52,53,106] | antioxidant, anticancer, antinociceptive, anticonvulsant, sedative, antibronchitis, antihypertensive, vasorelaxant, cardioprotective [107]; |
| 30 | 4-Terpineol | 🌱 | [49,53] | antimicrobial [108]; gastroprotective [109]; | |
| 31 | Thymol | monocyclic monoterpene hydrocarbon | 🌱✿ | [49,50] | anti-inflammatory, antioxidant, antimicrobial, immunostimulatory, anticancer [110]; cardioprotective [111]; antihypertensive [70]; antihyperglycemic [112]; antinociceptive [113]; gastroprotective [114]; anxiolytic [115]; |
| 32 | Abscisic acid | oxygenated monocyclic sesquiterpene | 🌱 | [116,117] | antidiabetic [118]; antinociceptive [119]; |
| 33 | α-Amorphene | polycyclic sesquiterpene hydrocarbon | 🌱 | [50,53] | n/a |
| 34 | Aromadendrene | polycyclic sesquiterpene hydrocarbon | 🌱✿ | [47,48,53] | antimicrobial [120]; |
| 35 | allo-Aromadendrene | polycyclic sesquiterpene hydrocarbon | 🌱✿ | [6,43,46,48,49,50,51,52,53] | |
| 36 | allo-Aromadendrene epoxide | oxygenated polycyclic sesquiterpene | 🌱 | [53] | |
| 37 | Bisabola-2,10-diene(1-9)oxide | oxygenated polycyclic sesquiterpene | 🌱 | [53] | n/a |
| 38 | β-Bisabolene | monocyclic sesquiterpene hydrocarbon | [48] | anticancer [121] | |
| 39 | epi-α-Bisabolol | monocyclic sesquiterpene alcohol | 🌱✿ | [46,48,49,53] | anti-inflammatory [122]; antimicrobial [123]; anticancer [124]; |
| 40 | α-Bisabolol | 🌱 | [43,53] | n/a | |
| 41 | β-Bisabolol | 🌱✿ | [48] | n/a | |
| 42 | β-Bourbonene | polycyclic sesquiterpene hydrocarbon | 🌱✿ | [6,43,46,47,49,50,51,52,53] | n/a |
| 43 | 1,5-di-epi-Bourbonene (α or β) | 🌱 | [53] | ||
| 44 | Bulnesol | polycyclic sesquiterpene alcohol | 🌱✿ | [49] | n/a |
| 45 | Cadalene | polycyclic sesquiterpene hydrocarbon | 🌱 | [53] | n/a |
| 46 | Cadina-1,4-diene | polycyclic sesquiterpene hydrocarbon | 🌱✿ | [48,53] | n/a |
| 47 | α-Cadinene | polycyclic sesquiterpene hydrocarbon | 🌱 | [50,53] | n/a |
| 48 | cis-γ-Cadinene | 🌱✿𐩕 | [49] | ||
| 49 | trans-γ-Cadinene | 🌱 | [46] | ||
| 50 | γ-Cadinene | 🌱 | [51,52,53] | ||
| 51 | δ-Cadinene | 🌱✿𐩕 | [43,46,48,49,50,52,53] | ||
| 52 | α-Cadinol | polycycylic sesquiterpene alcohol | 🌱✿ | [43,48,49,53] | antifungal [125]; |
| 53 | T-Cadinol | [46,49,53,126] | anticancer [127] | ||
| 54 | α-Calacorene | polycyclic sesquiterpene hydrocarbon | 🌱 | [50,53] | n/a |
| 55 | β-Calacorene | [53] | |||
| 56 | Calamenene | polycyclic sesquiterpene hydrocarbon | 🌱 | [50,53] | anticancer [127]; |
| 57 | Caryophylladienol I | polycyclic sesquiterpene alcohol | 🌱 | [53] | n/a |
| 58 | Caryophylladienol II | [53] | |||
| 59 | β-Caryophyllene | polycyclic sesquiterpene hydrocarbon | 🌱✿𐩕 | [6,43,46,47,48,49,50,51,52,53] | antioxidant, antimicrobial, antitumor, anticancer [128]; anti-inflammatory, neuroprotective [129]; anxiolytic, antidepressant [130]; anticonvulsant [131]; analgesic [132]; |
| 60 | β-Caryophyllene epoxide | oxygenated polycyclic sesquiterpene | 🌱 | [43,46,48,49,53] | anticancer, analgesic [132]; |
| 61 | Caryophyllenol II | polycyclic sesquiterpene alcohol | 🌱 | [53] | n/a |
| 62 | 8,14-Cedranoxide | oxygenated sesquiterpene | 🌱 | [46] | n/a |
| 63 | α-Cedrene | polycyclic sesquiterpene hydrocarbon | 🌱 | [47] | n/a |
| 64 | α-Copaene | polycyclic sesquiterpene hydrocarbon | 🌱 | [6,46,47,49,50,52,53] | antioxidant, anticancer [133]; neuroprotective [134]; |
| 65 | β-Copaene | 🌱✿𐩕 | [46,50,53] | ||
| 66 | α-Corocalene | polycyclic sesquiterpene hydrocarbon | 🌱 | [53] | n/a |
| 67 | α-Cubebene | polycyclic sesquiterpene hydrocarbon | 🌱✿ | [6,49,53] | antioxidant, neuroprotective [135]; antimicrobial [136]; anti-inflammatory [137]; |
| 68 | β-Cubebene | 🌱 | [6,46,49] | ||
| 69 | Cubebol | polycyclic sesquiterpene alcohol | 🌱 | [53] | n/a |
| 70 | 4-epi-Cubebol | [53] | |||
| 71 | 1,10-di-epiCubenol | polycyclic sesquiterpene alchohol | 🌱✿ | [46,49,53] | n/a |
| 72 | Cubenol | 🌱 | [53] | ||
| 73 | 1-epi-Cubenol | 🌱✿ | [46,49,53] | ||
| 74 | ar-Curcumen-15-al | oxygenated monocyclic sesquiterpene | 🌱 | [46,53] | n/a |
| 75 | ar-Curcumene | monocyclic sesquiterpene hydrocarbon | 🌱✿ | [6,43,46,47,48,49,50,51,52,53,126] | n/a |
| 76 | β-Curcumene | ✿ | [48] | ||
| 77 | γ-Curcumene | 🌱✿ | [48,49] | ||
| 78 | Curcuphenol | monocyclic sesquiterpene alcohol | 🌱✿ | [48,53] | anticancer [138]; |
| 79 | Cyclosativene | polycyclic sesquiterpene hydrocarbon | 🌱✿ | [49] | n/a |
| 80 | Dehydrosesquicineole | oxygenated polycyclic sesquiterpene | 🌱 | [53] | n/a |
| 81 | Bicyclo-Elemene | polycyclic sesquiterpene hydrocarbon | 🌱 | [50] | n/a |
| 82 | β-Elemene | monocyclic sesquiterpene hydrocarbon | 🌱 | [48,53] | anticancer, anti-inflammatory [139]; |
| 83 | γ-Elemene | 🌱 | [48] | ||
| 84 | δ-Elemene | 🌱✿ | [48,49,50] | ||
| 85 | Elemol | monocyclic sesquiterpene alcohol | 🌱 | [43,48,49] | n/a |
| 86 | β-Eudesma 4(15), 7 dien-1β-ol | polycyclic sesquiterpene alcohol | 🌱 | [53] | n/a |
| 87 | α-Eudesmol | 🌱 | [46] | neuroprotective [140]; | |
| 88 | β-Eudesmol | 🌱✿ | [46,48] | anti-allergic, anti-inflammatory [141]; anticancer [142]; | |
| 89 | γ-Eudesmol | 🌱✿ | [48,49] | n/a | |
| 90 | 10-epi-γ-Eudesmol | [48,49] | n/a | ||
| 91 | Kunseaol | monocyclic sesquiterpene alcohol | 🌱 | [53] | n/a |
| 92 | Bicyclo-Germacrene | polycyclic sesquiterpene hydrocarbon | 🌱 | [53] | n/a |
| 93 | Germacrene B | monocyclic sesquiterpene hydrocarbon | 🌱 | [47,48] | |
| 94 | Germacrene D | 🌱✿𐩕 | [6,43,47,48,49,51,52,53] | anticancer [143]; anti-inflammatory, analgesic [144]; antioxidant [145]; | |
| 95 | Iso-Germacrene D | 🌱 | [53] | ||
| 96 | β-Germacrenol | monocyclic sesquiterpene alcohol | 🌱 | [53] | n/a |
| 97 | Globulol | polycyclic sesquiterpene alcohol | 🌱✿ | [46,48] | n/a |
| 98 | α-Guaia-6,10(14)-diene-4β-ol | polycyclic sesquiterpene alcohol | 🌱 | [53] | n/a |
| 99 | Guaiene | polycyclic sesquiterpene hydrocarbon | 🌱 | [46] | n/a |
| 100 | Guaiol | polycyclic sesquiterpene alcohol | 🌱✿ | [48,49] | n/a |
| 101 | α-Gurjunene | polycyclic sesquiterpene hydrocarbon | 🌱✿ | [47,49,51,52] | n/a |
| 102 | β-Gurjunene | [49] | |||
| 103 | β-Himachalene | polycyclic sesquiterpene hydrocarbon | ✿𐩕 | [49] | n/a |
| 104 | α-Humulene | monocyclic sesquiterpene hydrocarbon | 🌱✿ | [6,43,46,47,48,49,51,52,53] | antitumor, anti- inflammatory, antimicrobial [146]; |
| 105 | Iso-Calamendiol | polycyclic sesquiterpene alcohol | 🌱 | [53] | n/a |
| 106 | Iso-Italicene | polycyclic sesquiterpene hydrocarbon | 🌱 | [46] | n/a |
| 107 | Juniper camphor | polycyclic sesquiterpene alcohol | 🌱✿ | [49] | n/a |
| 108 | Ledol | polycyclic sesquiterpene alcohol | 🌱 | [53,106] | n/a |
| 109 | α-Longipinene | polycyclic sesquiterpene hydrocarbon | 🌱 | [46] | n/a |
| 110 | cis-Muurola-4(14),5-diene | polycyclic sesquiterpene hydrocarbon | 🌱✿ | [49,53] | n/a |
| 111 | α-Muurolene | [6,48,49,50,52] | |||
| 112 | γ-Muurolene | [46,49,53] | |||
| 113 | 14-hydroxi-α-Muurolene | polycyclic sesquiterpene alcohol | 🌱 | [46] | n/a |
| 114 | α-Muurolol | 🌱✿ | [49] | ||
| 115 | epi-α-Muurolol | [48] | |||
| 116 | T-Muurolol | [43,46,49,49,53] | |||
| 117 | E-Nerolidol | acyclic sesquiterpene alcohol | 🌱✿ | [48] | antihyperlipidemic, anti-inflammatory, anti-uterine fibroids [147]; anticancer [148]; |
| 118 | (E)-Nuciferol | moncyclic sesquiterpene alcohol | 🌱 | [53] | n/a |
| 119 | β-Oplopenone | oxygenated polycyclic sesquiterpene | 🌱 | [53] | n/a |
| 120 | Salvial-4(14)-en-1-one | oxygenated polycyclic sesquiterpene | 🌱 | [53] | n/a |
| 121 | α-Santalene | polycyclic sesquiterpene hydrocarbon | 🌱 | [50] | n/a |
| 122 | cis-α-Santalol | polycyclic sesquiterpene alcohol | 🌱 | [46] | antihyperglycemic, antioxidant [149]; |
| 123 | Selin-11-en-4-α-ol | polycyclic sesquiterpene alcohol | 🌱 | [106] | anxiolytic, sedative [150]; |
| 124 | Selina-3,7(11)-diene | polycyclic sesquiterpene hydrocarbon | 🌱 | [50] | n/a |
| 125 | α-Selinene | polycyclic sesquiterpene hydrocarbon | 🌱 | [50] | n/a |
| 126 | β-Sesquiphellandrene | monocyclic sesquiterpene hydrocarbon | 🌱✿ | [43,47,49,53] | anticancer [151]; antioxidant [152]; |
| 127 | trans-Sesquisabinene hydrate | polycyclic sesquiterpene alcohol | 🌱 | [48] | n/a |
| 128 | Shyobunone | oxygenated monocyclic sesquiterpene | 🌱 | [6,53] | neuroprotective, acetyl-cholinesterase inhibition [153]; |
| 129 | 6-epi-Shyobunone | [53] | |||
| 130 | iso-Shyobunone | [53] | |||
| 131 | Spathulenol | polycyclic sesquiterpene alcohol | 🌱✿ | [46,48,49,53] | neuroprotective [154]; antibacterial, antioxidant, anti-inflammatory, anticancer [155]; |
| 132 | Spathulenol isomer | 🌱 | [48] | ||
| 133 | ar-Turmerol | monocyclic sesquiterpene alcohol | 🌱✿ | [46,48] | n/a |
| 134 | Valerianol | polycyclic sesquiterpene alcohol | 🌱✿ | [49] | n/a |
| 135 | Viridiflorol | polycyclic sesquiterpene alcohol | 🌱✿ | [48,53] | anti-arthritic, analgesic, antinociceptive [156]; anticancaer [157]; antioxidant, antibacterial, anti-inflammatory [158]; |
| 136 | Xanthorrhizol | monocyclic sesquiterpene alcohol | 🌱 | [46,53] | anticancer [159]; antimicrobial, antibacterial [160]; antihypolipidemic [161]; anti-inflammatory [162]; |
| 137 | α-Ylangene | polycyclic sesquiterpene hydrocarbon | 🌱 | [50] | n/a |
| 138 | β-Ylangene | 🌱 | [53] | ||
| 139 | α-Zingiberene | monocyclic sesquiterpene hydrocarbon | 🌱✿𐩕 | [6,43,46,47,48,49,50,51,52,53] | analgesic, neuroprotective [163]; anticancer [164]; anti-inflammatory [165]; |
| 140 | Zingiberenol | monocyclic sesquiterpene alcohol | 🌱 | [53] | n/a |
| 141 | 15,16-Dinorlabd-8(20)-en-13-one | oxygenated diterpene | 🌱 | [106] | n/a |
| 142 | Geranyl-p-cymene | monocyclic diterpene hydrocarbon | 🌱 | [53] | n/a |
| 143 | Geranyl α-terpinene | monocyclic diterpene hydrocarbon | 🌱 | [53] | n/a |
| 144 | Geranyl linalool | acyclic diterpene alcohol | 🌱 | [53] | n/a |
| 145 | Gibberellin | oxygenated polycyclic diterpene | 🌱 | [29] | n/a |
| 146 | Manool | polycyclic diterpene alcohol | 🌱 | [106] | antioxidant, anti-inflammatory [166]; anticancer [167]; antihypertensive [168]; cardioprotective [169]; |
| 147 | Manoyl oxide | polycyclic oxygenated diterpene | 🌱 | [126] | anticancer [170]; |
| 148 | 13-epi-Manoyl oxide | [47,53,126] | |||
| 149 | Methyl Neoabietate | oxygenated polycyclic diterpene | 🌱 | [47] | n/a |
| 150 | lutein | oxygenated polycyclic tetraterpene | 🌱 | [29] | antioxidant, anti-inflammatory, neuroprotective [171]; anticancer [172]; hepatoprotective [173]; cardiopreotective [174]; |
| 151 | neoxanthin | oxygenated polycyclic tetraterpene | 🌱 | [29] | antihyperlipidemic [175]; |
| 152 | zeaxanthin | oxygenated polycyclic tetraterpene | 🌱 | [29] | n/a |
The following quantities refer to the w/w of the extracted oil. Oxygenated sesquiterpenes ranged from 44.8% in aerial parts (twigs, leaves, flowers) [46] to 67.1% in leaves [49]. Hydrocarbon sesquiterpenes were found from 22.5% in flowers [49] to 48.6% in aerial parts including flowers [46]. Within representative sesquiterpenes in leaves, α-zingiberene is the most abundant and was extracted from 5.9% [49] to 14.8% [48], followed by α-bisabolol with values ranging from 1.9% [48] to 11.4% [46]. Further ar-curcumene was obtained from 8.3% [46] to 10.6% [48], and β-bourbonene showed a presence from a residual 0.1% [48] up to 8.7% [46].
The essential oil compositions of the aerial parts (stems and leaves) of C. albidus showed only quantitative differences. However, the flowers’ (petals’) essential oil has a different composition, with mainly α-zingiberene, α-cadinol, ar-curcumene, and δ-cadinene [48,49], while the composition of the isolated pollen contained α-zingiberene, δ-cadinene, and germacrene D within the most abundant compounds [49].
Since the analyzed samples come from different places with different climatic and soil conditions, were collected on different dates, and were sometimes analyzed by different methods, comparison of their compositions is only possible to a very limited extent. In addition to these limitations, it must also be taken into account that the species of the Cistus subgenus hybridize with each other, which may also have an impact on the composition of the synthesized compounds if species purity has not been ensured beforehand.
5.1.2. Phenylpropanoids from the Essential Oils
Phenylpropanoids are compounds that are built from an aromatic benzene ring and a chain of three carbon atoms and often have hydroxyl and methoxy groups on the aromatic ring. Many phenylpropanoids are natural substances that are formed in plants and microorganisms through the shikimate biosynthetic pathway, with phenylalanine and tyrosine as intermediate compounds [176]. In addition to terpenes, phenylpropanoids are a frequent component of essential oils and represent the majority of natural phenolic substances or their precursors. The phenolpropanoids identified so far in C. albidus are eugenol [49,53,177] and chavicol [50].
5.1.3. Diterpenes
As for the diterpenes present in C. albidus, these are not usually detected in most analyses, because they belong to the non-volatile terpenes. This is mainly because the analytical methods described are not suitable for detecting non-volatile substances and not because of the absence of these compounds. The diterpenes detected so far are geranyl linalool, geranyl α-terpinene, geranyl p-cymene and 13-epi-mannoyloxide [53], methyl neoabietate [47], 15,16-dinorlabd-8(20)-en-13-one, manool [106], and manoyl oxide [126]. The latter was found in the aerial parts, including the stems, unlike 15,16-dinorlabd-8(20)-en-13-one and manool, which were obtained only from the leaves.
5.1.4. Tetraterpenes
Tetraterpenes are vital for plant growth, protection against stress, and successful reproduction. So far, lutein, neoxanthin, and zeaxanthin have been identified in C. albidus [29].
5.2. Phenolic Compounds
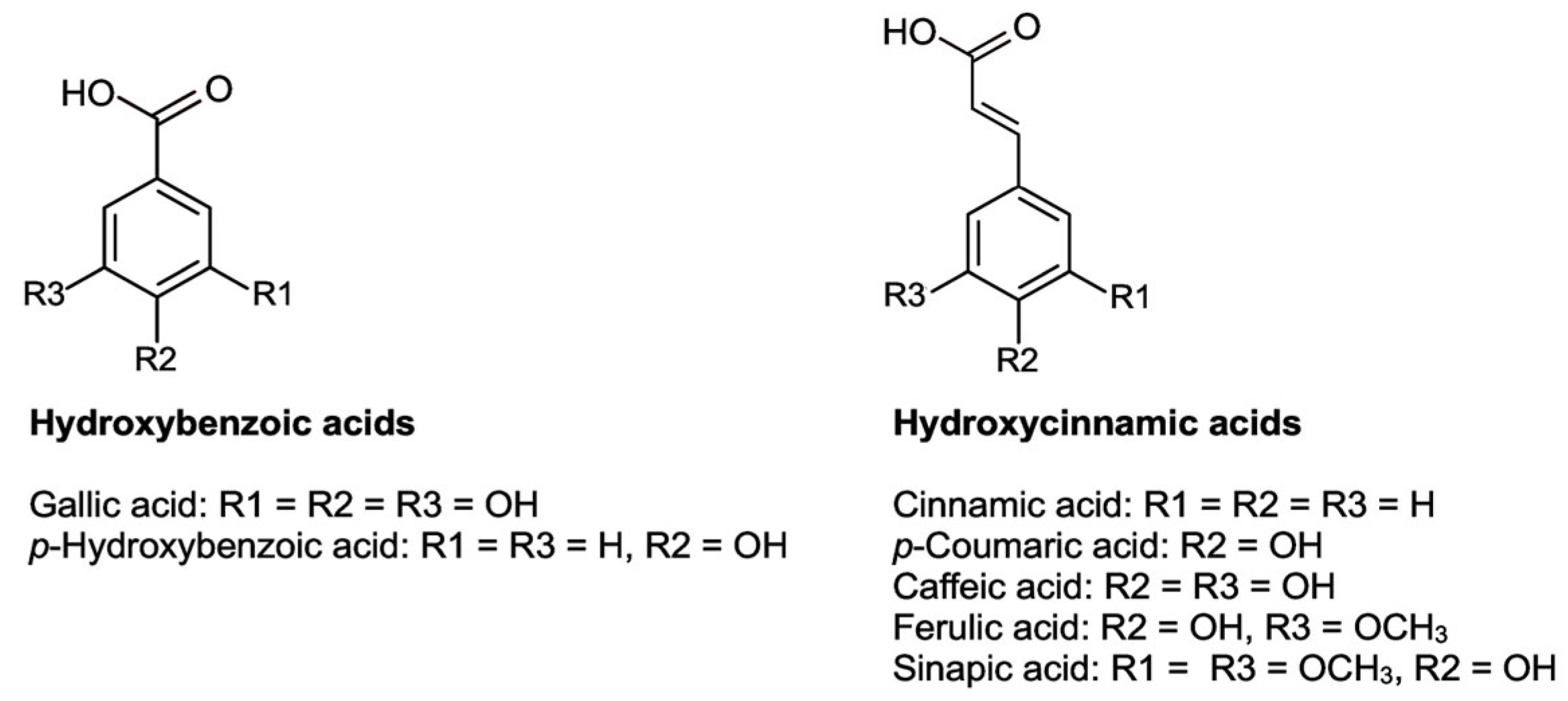
Phenolic compounds are based on the phenol structure. In general, these compounds can be divided into seven subgroups: simple phenols, hydroxybenzoic acids, hydroxycinnamic acids, coumarins, flavonoids, lignans, and lignins [178]. The concentration of these phenolic substances in plant foods depends, in part, on the plant species, the climate, and the degree of maturity [179]. In the present work, the analytical focus of phenolic compounds lies on the group of hydroxybenzoic acids (basic structure C6-C1), hydroxycinnamic acids (basic structure C6-C3) (Figure 5), and flavonoids (basic structure C6-C3-C6) as they are the most frequent polyphenolic groups present in C. albidus samples (Figure 6).
In general terms, the polyphenolic composition of C. albidus, listed in Table 2, is very similar to that of other members of the Cistus subgenus, such as C. crispus and C. × incanus (hybrid of C. albidus × C. criticus) [5]. A semi-quantitative analysis of the composition of extracts from C. albidus, C. clusii, C. ladanifer, and C. salviifolius revealed small differences between them [177] and a series of substances that occur exclusively in C. albidus, namely caftaric acid, prunin, and 5-O-caffeoylquinic acid glucoside [12].
5.2.1. Flavonoids
Flavonoids contribute to overall fruit color and flavor in plants [178]. In the form of flavones, they are responsible for the yellow hues of the inner petals in the flowers and of the stamens and, in the form of anthocyanidins, for the purple-pink colored petals of C. albidus. Flavones often appear as co-pigments of anthocyanins. The interaction of both types of dye explains the simultaneous appearance of yellow and red in different flowers. The flavonoids found so far are flavonols, flavones, flavanols, and tannins (Figure 7).
Flavonols and flavones occur in C. albidus’ aerial parts as free algycones and glycosides. They are responsible for yellow color nuances and are usually tasteless. Flavonols are distinguished from flavones by the presence of a 3-hydroxyl group (Figure 7). Phenolic substances of the flavonol subclass are present in all plant organs of C. albidus. Characteristic representatives of these compounds in C. albidus are kaempferol, quercetin, myricetin, myricitrin (myricetin-3-O-rhamnoside), and quercitrin (quercetin-3-O-rhamnoside) [5]. High contents of myricetin glycoside (7 mg/g dry weight) were detected in C. albidus aerial parts, with myricetin-3-O-rhamnoside being the primary derivative of myricetin with 83% of total myricetin composition, followed by quercetin glycoside (2 mg/g dry weight), with quercetin-3-O-rhamnoside being the most abundant derivative of quercetin, constituting 80% of the total quercetin composition [11]. The only flavones detected so far are diglycosylated apigenin [5] and the isoflavone glycitin 6″-O-malonate [12].
Flavanols are found predominantly in the leaves of C. albidus and contribute particularly to the astringent taste of extracts. Compounds belonging to this subclass have two asymmetrically substituted carbon atoms and can therefore exist as diastereomeric 5,7,3-,4-tetrahydroxyflavanols, catechin and epicatechin. A third hydroxyl group on ring B also results in 5,7,3,4,5-pentahydroxyflavanol gallocatechin, or correspondingly epigallocatechin (Figure 7). Among the polymeric flavanols, the most relevant compounds in C. albidus are prodelphinidins such as (epi-)gallocatechins. Procyanidins are other representative flavanols contained in C. albidus leaves. Within these oligomeric compounds, (epi-)catechins are most common in C. albidus aerial parts [5]. However, combinations of prodelphinidins and procyanidins have also been detected. So far, the following flavan-3-ol compounds have been found: (+)-catechin, (−)-epicatechin, (−)-(epi)gallocatechin, (−)-(epi)gallocatechin-(epi)catechin dimer, (−)-(epi)gallocatechin, and (−)-(epi)gallocatechin-(epi)gallocatein dimer [5,180].
Tannins are water-soluble, slightly acidic oligomers of polyphenols. They are able to form water-insoluble complexes with protein molecules. In the past, this property of C. albidus was used to tan animal skin in leather production [181]. Based on the chemical structure of the monomeric building blocks, tannins present in C. albidus could be divided into two groups, the condensed and the hydrolyzable tannins.
The first group includes the proanthocyanidins (flavanols) already described, which are also known as condensed tannins. They consist of polymerized flavonoid phenols, such as catechins, epicatechin, anthocyanins, and so on. They are correspondingly polymers whose monomeric units consist of phenolic flavans, mostly catechin (flavan-3-ol).
The second group represents hydrolyzable tannins, which are hydrolyzed by the action of acids. These compounds exist as various polyhydroxy compounds, for example as sugar esterified with a phenolic acid [178]. Within this group, hexahydroxydiphenoyl-D-glucose (HHDP-Glc) was detected in C. albidus [5]. When the phenolic acid is gallic acid, the compound is called gallotannin. If, on the other hand, esterification with hexahydroxydiphenic acid occurs, this compound is called ellagitannin. Ellagitannins were not found in C. albidus aerial parts, except for a residual presence of glucogallin, pedunculagin, and punicalagin gallate [5,11].
5.2.2. Phenolic Acids
Phenolic acids are understood as the hydroxylated derivatives of benzoic acids (hydroxybenzoic acids) and cinnamic acids (hydroxycinnamic acids). Their highest concentration in plants is found in the outer leaves. In C. albidus aerial parts, glucogallin [5,11] and gallic acid [5,182] were found. Within the category of phenolic acid derivatives, 5-O-caffeoyl quinic acid glucoside, caffeoylquinic glycoside [12], and uralenneoside and rhamnoside of hydroxyferulic acid [5] have also been detected.
Table 2.
Phenolic compounds identified in Cistus albidus. 🌱: aerial parts, including leaves and twigs. n/a: reliable data are not available.
Table 2.
Phenolic compounds identified in Cistus albidus. 🌱: aerial parts, including leaves and twigs. n/a: reliable data are not available.
| No | Compound | Structure/Class | Presence in | Analytical Reference | Pharmacology |
|---|---|---|---|---|---|
| 1 | Apigenin diglucoside | flavone | 🌱 | [5] | anticancer, anti-inflammatory, antimicrobial, antioxidant [183]; anxiolytic [184]; |
| 2 | Caffeic acid | phenolic acid | 🌱 | [11] | anticancer [185,186,187]; antimutagenic, antihyperglycemic, anti-inflammatory, antioxidant, [188,189]; antmicrobial [186]; cardioprotective [189]; hepatoprotective [190]; neuroprotective [187,191]; |
| 3 | Caftaric acid | phenolic acid | 🌱 | [12] | anticancer, antidiabetic, antihypertensive, anti-inflammatory, antimutagenic, antioxidant, hepatoprotective, [192]; |
| 4 | Cynarin | phenolic acid | 🌱 | [12] | anticancer, antidiabetic antiulcer, antivirus, antioxidant, hepatoprotective, hypocholesterolemic [193]; cardioprotective [194]; |
| 5 | (-)-(Epi)catechin | flavanol | 🌱 | [5,177,180] | neuroprotective [195]; antioxidant [195,196]; antimicrobial [197]; anti-inflammatory, antiallergenic, antivirus, anticancer, skin penetration enhancer, UV-protection [198]; |
| 6 | (-)-(Epi)gallocatechin | 🌱 | [5,177,180] | ||
| 7 | Epigallocatechin-(4β-->8)-gallocatechin-(4α-->8)-gallocatechin trimer | 🌱 | [5,180] | ||
| 8 | Epigallocatechin-(4β-->8)-gallocatechin-(4α-->8)-catechin trimer | 🌱 | [5,180] | ||
| 9 | (-)-(Epi)catechin-(epi)gallocatechin dimer | 🌱 | [5] | ||
| 10 | (-)-(Epi)gallocatechin-(epigallocatechin dimer | 🌱 | [5] | ||
| 11 | (-)-(Epi)gallocatechin gallate | 🌱 | [5] | ||
| 12 | 4,3′,4′-Trimethyl catechin | 🌱 | [12] | ||
| 13 | Ferulic acid oligomer | phenolic acid | 🌱 | [12] | antioxidant [199]; antimicrobial [200]; anti-inflammatory [201]; neuroprotective [202]; antiviral [203]; antiallergic [204]; hepatoprotective [205]; anticancer [206]; antithrombotic [207]; antidiabetic [208]; |
| 14 | Hydroxy-ferulic acid rhamnoside | phenolic acid glycoside | 🌱 | [5] | |
| 15 | Feruoyl quinic glucoside | 🌱 | [12] | n/a | |
| 16 | Gallic acid | phenolic acid | 🌱 | [5,182] | anti-inflammatory [209,210]; antiobesity, antioxidant [211]; hepatoprotective [210]; anticancer [212]; antifungal [213]; |
| 17 | Galloyl glucose | flavanol | 🌱 | [12] | antiviral [214]; |
| 18 | Glucogallin | flavanol | 🌱 | [5,11] | antioxidant, anti-inflammatory, antidiabetic, cataract-preventing, antiglaucoma, UV-protective [215]; |
| 19 | Glycitin 6″-O-malonate | isoflavone (flavonoid glycoside) | 🌱 | [12] | n/a |
| 20 | Hexahydroxydiphenoyl- D-glucose | hydrolysable tannin | [5] | antioxidant [216] | |
| 21 | Kaempferol diglucoside | flavonol | 🌱 | [5] | antidiabetic [217]; anxiolytic, antidepressant, antiepileptic, anti-inflammatory, neuroprotective, analgesic [218]; |
| 22 | Kaempferol 3-O-rutinoside | 🌱 | [5,177] | ||
| 23 | Luteolin-7-O-rutinoside | flavonol | 🌱 | [12] | antioxidant, anti-inflammatory [219]; |
| 24 | Myricetin hexoside | flavonol | 🌱 | [5,177] | anticancer, antidiabetic, antiobesity, cardioprotective, osteoporosis protective, anti-inflammatory, hepatoprotective [220]; |
| 25 | Myricetin glycoside | 🌱 | [11] | ||
| 26 | Myricetin 3-O-rutinoside | 🌱 | [12] | ||
| 27 | Myricetin-O-galloyl-hexoside | 🌱 | [12] | ||
| 28 | Myricitrin | 🌱 | [5] | ||
| 29 | Oenin | anthocyanin (flavonoid glycoside) | 🌱 | [12] | anticancer [221]; neuroprotective [222]; anti-inflammatory [223]; antioxidant [224]; |
| 30 | Pedunculagin | ellagitannin | 🌱 | [5,11] | antioxidant, anti-inflammatory, dermatoprotective [225]; anticancer [226]; antibacterial [227]; |
| 31 | Pelargonidin 3-O-(6″malonylglucoside) | anthocyanin (flavonoid glycoside) | 🌱 | [12] | cardioprotective, neuroprotective [228,229]; |
| 32 | Peonidin 3-O-(6″-p-coumaroyl) glucoside | anthocyanin (flavonoid glycoside) | 🌱 | [12] | cardioprotective, neuroprotective [228,229]; |
| 33 | Petunidin | anthocyanidin (O-methylated flavonoid) | 🌱 | [12] | antioxidant [230]; cardioprotective, neuroprotective [228,229]; |
| 34 | Procyanidin | flavanol | 🌱 | [12] | antioxidant [231]; antibacterial [232]; anticancer [233]; anti-inflammatory [231,234]; |
| 35 | Prunin | flavanone | 🌱 | [12] | antihyperlipidemic, antihyperglycemic, antidiabetic [235]; |
| 36 | Punicalagin | ellagitannin | 🌱 | [11] | anti-inflammatory, antioxidant, neuroprotective, hepatoprotective, cardioprotective, antiviral, antimicrobial, anticancer, antidiabetic, antihyperlipidemic, gastroprotective [236]; |
| 37 | Punicalagin gallate | 🌱 | [11] | ||
| 38 | Quercetin | flavonol | 🌱 | [12] | anticancer, antitumor, anti-inflammatory, antiviral, antihypercholesterolemia, antihyperglycemic, antioxidant, antibacterial, cardioprotective, gastroprotective, hepatoprotective, antihypertensive, nephroprotective, neuroprotective [237]; antidepressant [238]; antimicrobial [239], antifungal [240]; antiallergic [241] antiobesity [242]; |
| 39 | Quercetin glucoside | 🌱 | |||
| 40 | Quercetin 3-O-glucoside | 🌱 | [12] | ||
| 41 | Quercetin 3-O-(2’-cumaroyl)- rutinoside | 🌱 | [12] | ||
| 42 | Quercetin 3,4-diglucoside | 🌱 | [12] | ||
| 43 | Quercetin 3-O-(2’-caffeoyl)-rutinoside | 🌱 | [5] | ||
| 44 | Quercitrin | 🌱 | [5,11] | ||
| 45 | Rutin | 🌱 | [5,11,12,177] | ||
| 46 | Quinic acid | phenolic acid | 🌱 | [12,177] | radioprotective [243]; neuroprotective [244]; anti-inflammatory [245]; 5antiviral [246]; |
| 47 | 3p-Coumaroylquinic acid | 🌱 | [12] | n/a | |
| 48 | 3-Caffeoylquinic acid | 🌱 | [11] | antioxidant, anti-inflammatory [247]; | |
| 49 | Caffeoylquinic glycoside | phenolic acid glycoside | 🌱 | [12] | enzyme inhibition, hepatoprotective [248]; |
| 50 | Isorhamnetin- O-rutinoside | flavonol | 🌱 | [5] | anti-atherosclerosis, cardioprotective, neuroprotective, anticancer, antihypertensive, antioxidant antihyperglycemic, hepatoprotective, anti-inflammatory, anti-osteoporosis, antiobesity, UV-protection [249]; antimicrobial [250]; |
| 51 | 3-O-Methylrosmarinic acid | caffeic acid ester | 🌱 | [12] | anti-inflammatory, antioxidant, antidiabetes, antivirus, antitumor, neuroprotective, hepatoprotective [251]; |
| 52 | Methoxy dihydroferuoyl methyl rosmarinic acid | 🌱 | [12] | ||
| 53 | Dihydroxy-dihydro feruoyl methyl rosmarinic acid | 🌱 | [12] | ||
| 54 | p-Hydroxy benzil rosmarinic acid | 🌱 | [12] | ||
| 55 | Shikimic acid dimer | phenolic acid | 🌱 | [12] | anticoagulant [252]; antithrombotic [253]; |
| 56 | 6′-O-Sinapoyl sucrose | hydroxy-cinnamate sucrose esters | 🌱 | [12] | antioxidant [254]; |
| 57 | Syringyl shikimic acid dimer | phenolic acid | 🌱 | [12] | n/a |
| 58 | Uralenneoside | p-hydroxy- benzoic acid alkyl ester | 🌱 | [5] | n/a |
5.3. Carbonylic Compounds
Aldehydes are odoriferous aromatic substances in plants. These often arise from substances containing linolenic acid during harvesting, crushing, or preparation [255]. The aliphatic aldehydes, octanal [49], nonanal, and decanal [49,53], were exclusively identified in pollen and may be responsible for the typical sweet smell of the flowers [256]. On the other hand, tetradecanal, undecanal, and dodecanal were present in the aerial parts [53]. These compounds may contribute to the typical sweetish odor of C. albidus with its undertone of oranges, lemons, and roses [256,257].
5.4. Phytohormones and Vitamin E
Tocopherols and tocotrienols are also present in leaves and seeds. More than 75% of the vitamin E present in the seeds was in the form of α-tocopherol, followed by α-tocotrienol and γ-tocopherol [29,35,117], which is the immediate precursor of α-tocopherol. Phytohormone and vitamin studies further revealed the presence of the jasmonates 12-oxo-phytodienoic acid (OPDA), jasmonic acid (JA), and jasmonoyl-isoleucine (JA-Ile) and plastochormanol-8 [20], carotenoids, and abscisic acid, with α-tocopherol being the most abundant [27]. A negative correlation was revealed between vitamin E and OPDA accumulation in C. albidus under winter conditions, while a positive correlation was found between JA and α-tocopherol [20]. A significant positive correlation was also detected between hydration and total leaf chlorophylls due to the protection mechanism of tocopherols from the photosynthetic apparatus. Therefore, higher levels of α-tocopherol were observed under abiotic stress conditions and when the leaves showed an orientation more perpendicular to the solar rays [20,27,28,258].
Drought stress can induce an increase in the concentrations of abscisic acid and H2O2 in the leaves, inducing an increase in ascorbic acid, maintaining and even decreasing the oxidative state of ascorbate, thus protecting plants from oxidative damage [30,259]. In addition, cytokinins that act as nitric oxide scavengers and are involved in the modulation of the abscisic acid response have been reported [29].
C. albidus seeds obtained from mature plants showed higher concentrations of α-tocopherol, JA, and salicylic acid than those obtained from younger plants. Auxin (indole-3-acetic acid) content was also significantly higher in seeds from older plants. Gibberellic acid GA4 and its precursor GA24 were also found in seeds [27]. No differences were detected between the concentration of cytokinins in seeds from older and younger plants, except for zeatin, which was significantly higher in seeds from older plants. Zeatin was the main form of cytokinin found in the seeds of C. albidus [27].
5.5. Alkanes
Among the volatile compounds isolated from aerial parts of C. albidus, tricosan [53], tetracosan [47], pentacosan [43], octacosan [47], and docosan [43,47] were found. n-Tridecane was present in petals, and n-tetradecane was identified in both petals and pollen, while n-hexadecane was detected only in pollen, and n-pentadecane was found in leaves [43,47,53].
5.6. Other Compounds: Fatty and Carboxylic Acids
Although fatty acids are products of the primary metabolism, they are described here for the role they may play in the bioavailability of pharmacologically active compounds, as described below. Among the fatty acids found in the aerial parts of C. albidus are tetradecanoic, primaric and pentadecanoic acid [47], nonanoic acid [106], palmitic acid [53], and butanoic acid [49]. The fatty acid composition of the seeds showed significantly higher levels of polyunsaturated fatty acids as well as very-long-chain saturated fatty acids for older plants due to their higher levels of linoleic acid [27]. In addition, the carboxylic acids methacrylic acid [106] and quinic acid [12,177] were identified.
6. Preparation Methods of the C. albidus Extracts
Studies of the pharmacological properties of traditional medicines based on C. albidus preparations are related to the presence of terpenoids and polyphenols. In order to understand the use that has been given to C. albidus traditionally, it is necessary to first review the preparation methods used in popular medicine of this plant due to their influence on the pharmacological effect.
6.1. Traditional Preparations
For the different applications, traditionally, only the aerial parts of C. albidus were harvested, mainly the leaves, but also flowers, flower buds, and to a lesser extent stems. The traditional preparation of C. albidus varies from an infusion to a prolonged decoction, while the dose usually used is around 3 g per 100 mL of water, taking a cup (150 mL) two or three times a day [10].
Within traditional preparations, decoction is the most used technique. It consists of boiling the plant material for a certain period of time and letting it rest afterwards. This method is primarily suitable for thermostable and water-soluble phytochemicals. During decoction, several compounds undergo chemical modifications. For example, catechins undergo epimerization, which is a change in their configuration relative to one of their stereogenic centres. Epimers, specifically epicatechins and epigallocatechins, have been shown to have important health benefits. It has been found that this epimerization occurs more readily in water with alkaline pH values than in purified water [260]. In addition, it has been shown that at temperatures greater than 98 °C, epimerization occurs faster than its degradation [261], so it can be deduced that the traditional preparation of C. albidus is the most effective way to extract catechins and their epimers. However, for green tea, the levels of epicatechin, epicatechin gallate, epigallocatechin, and epigallocatechin gallate were reported to increase only during the first 3 to 5 min of preparation (infusion at 85 °C), and the proportion of these flavonoids decreased as time increased. In contrast, another study found that levels of catechin, gallocatechin, and gallocatechin gallate increased continuously with the length of preparation time [262]. Taking these results into account, the pharmacological activities referred to in traditional use could be optimized by limiting the decoction time. Nonetheless, thermolabile compounds are lost in the decoction process. As a result, monoterpenes should not be contained in the resulting extract. Sesquiterpenes, however, would not be affected by extracting temperatures around 100 degrees but by low solubility in water due to their lipophilic character. It can therefore be assumed that terpenes play a minor role in the traditional decoction of plant material.
On the other side, probably in order to use the entire compound spectrum of the plant, based on both terpenes and polyphenols, the dried and crushed leaves sometimes were used directly (orally) [9,263,264,265,266]. This usage ensures that the resulting medicine is rich in polyphenols terpenes and other volatiles. However, a loss of several terpenes and oxidative reactions could be induced by the drying process, as reported for other species as Cannabis sativa [267,268].
6.2. Actual and Alternative Extraction Methods
In general, solid liquid extraction is the most commonly used method for extractive purposes. When using water as a solvent, the extraction is carried out at temperatures ranging between 20 and 50 °C. For extraction with solvents such as ethanol, methanol, and acetone, temperatures commonly used range from 60 to 80 °C [269]. For temperatures above 80 °C, a decrease in the yield of total polyphenols and proanthocyanidins has been reported, suggesting that it is due to their degradation; this process accelerates with temperatures above 70–80 °C [270,271]. However, other studies have shown that the higher the temperature, the more efficient the extraction is, since the heat makes the cell walls permeable, increasing the solubility and the diffusion coefficients of the compounds to be extracted and decreasing the viscosity of the solvent [272]. The extraction temperature has a significant effect on the types of polyphenols that are extracted. However, the thermal degradation of different types of polyphenols varies according to the pre-treatment, type of solvent, pH, and time used [269].
On the other hand, a mixture of polar and nonpolar solvents should result in an efficient extraction of both polyphenols and terpenes at the same time. However, with a few exceptions, in its traditional use, C. albidus has not been extracted by solvents with combined properties, such as EtOH. Studies have revealed low efficiency of these extraction methods, as well as environmental problems due to the requirement of large volumes of solvents, especially if they are organic [273]. Furthermore, traditional extraction processes often involve an intermediate step before extract concentration, which is a time- and energy-consuming process.
The compounds of C. albidus listed in this review were obtained from the aerial parts of the plant by means of distillation or extraction (water, ethanol, chloroform, pentene). Essential oils were isolated from fresh aerial parts by hydro-distillation using a Clevenger’s apparatus for between 2 h and 8 h. The isolated oil was dried over anhydrous sodium sulphate, and the essential oils were stored at +4 °C, in glass vials at dark, until analyses [46,48,49,126]. Pentane extracts [47] and a mixture of pentane/diethyl oxide [53] were also used to isolate essential oils.
To analyze the polyphenols of C. albidus, aerial parts were usually air dried and ground into powder to a size between 2 and 5 mm and then extracted by ethanol maceration (1:4, w/v) up to 72 h or acidified (0.1% HCl) methanol (1:3, w/v) for 24 h. The plant extracts were filtered or decanted (<24 h) and dried under vacuum using a rotary evaporator at temperatures ≤ 50 °C. Extracts were stored in the dark between 2 and 8 °C until used for the assessment [12,177,274].
A special case of polyphenol extraction is the flavonol-enriched C. albidus chloroform extract described by Tahiri et al. [13]. Here, the supernatant of the ethanol extract was further partitioned in ethyl acetate and water (1:3:1, w/v/v) and then separated into an organic and an aqueous phase. The organic phase of ethyl acetate was further partitioned in chloroform and water (1:3:1, w/v/v), yielding an organic and an aqueous phase of chloroform, respectively.
For the aqueous extracts, fresh aerial parts were ground to a maximum size of 5 mm and macerated for 2 h with distilled water (1:5–20, w/v) at temperatures between 60 and 90 °C. The filtered extract was then centrifuged and concentrated by rotatory evaporation and kept at 4 °C until use [5,11,177,182].
Alternative methods include supercritical CO2 extraction, microwave-assisted extraction, ultrasound-assisted extraction, enzyme-assisted extraction, pressurized fluid extraction, or a combination of these approaches. These methods require less solvent volume and extraction time, while at the same time producing higher yields and reducing toxic residues. Although these methods have not yet been described with C. albidus, they may be an interesting alternative that should be further explored, as has been done for other vegetal matrices from Rosmarinus officinalis [275] and Salvia miltiorrhiza [276].
7. Bioavailability of the Groups of Compounds Found in the C. albidus Extracts
Bioavailability is a pharmacokinetic concept that refers to the proportion and the pace at which a served dose of a drug reaches its therapeutic target, considering the tissue on which it acts. According to this definition, this term includes the absorption, metabolism, and distribution of a single compound. In general, secondary plant substances are absorbed, distributed, and metabolized in a similar way to xenobiotics [176].
Despite the fact that it has not yet been proven that extracts from C. albidus will be bioavailable, in this subsection, an attempt will be made to derive the bioavailability theoretically.
The structure of flavonoids affects their solubility in water, which in turn affects their absorption and bioavailability. Flavonoids with a more hydrophobic structure. such as flavonols, may have lower bioavailability compared to those that are more hydrophilic, such as flavanols. Flavonols contain a high proportion of hydrophobic groups, such as aromatic rings and aliphatic chains, which make them less soluble in water and more soluble in lipids [178]. Flavonoids with a more hydrophobic structure present in C. albidus are apigenin, quercetin, myricetin, and kaempferol. They must undergo metabolism in the gut to be absorbed [277].
On the other hand, the bioavailability of flavanols and condensed tannins is related to their catechol ring structure with hydroxyl groups that can form hydrogen bonds with water molecules, which makes them more soluble in water and more readily absorbed by the body. They cannot be hydrolyzed in the stomach and are therefore broken down into phenolic acids by microorganisms in the large intestine. The latter are then resorbed in the large intestine [277]. Thus, flavanols, such as catechins, are relatively soluble in water and can be absorbed by the body through passive diffusion in the small intestine [278].
The gut microbiota plays a crucial role in the metabolism and absorption of flavonoids by breaking them down into smaller metabolites that can be absorbed and utilized by the body. Flavonoids are not readily absorbed in their native form and must therefore undergo extensive metabolism in the gut to be absorbed and utilized [277]. The gut microbiota contains a diverse community of microorganisms, including bacteria, fungi, and viruses, that can metabolize flavonoids through various pathways. Some bacterial species have specialized enzymes that can break down flavonoids into smaller metabolites that are more bioavailable, such as phenolic acids and aromatic compounds [278,279].
Phenolic acids are generally more water-soluble than flavonoids and can be absorbed by the body through passive diffusion in the small intestine. Overall, the bioavailability of unesterified phenolic acids is high. These can be absorbed quickly in the small intestine without prior hydrolysis. However, phenolic acids are mainly present in esterified form, so that due to the lack of esterases in the human digestive tract, there is no absorption in the small intestine. Esterified phenolic acids are therefore first hydrolyzed by microorganisms in the large intestine, and the resulting metabolites are then resorbed [278].
Ellagitannins found in C. albidus are considered to have relatively low bioavailability. This is because they are highly complex molecules that require microbial metabolism in the colon to be broken down into smaller, absorbable metabolites [278].
Regarding the terpenes and other poorly absorbed compounds, it is said that both their low permeability across absorption barriers and reduced solubility in biological fluids decreases their bioavailability [178]. However, it might be increased through the natural presence in C. albidus of certain unsaturated fatty acids by enhancing micellarization during the digestion process. [280]. These compounds might increase the absorption of terpenoids and polyphenols present in C. albidus samples.
Furthermore, phenolic and terpenoid compounds, in the presence of α-tocopherol, have been shown to promote transport through the blood brain barrier (BBB) [281]. Moreover, studies have shown that borneol could increase the permeability of BBB by significantly losing the intercellular tight junctions (TJ) and increasing the number and volume of fluid endocytosis (pinocytosis) in in vitro models, thus enhancing the bioavailability of drugs [282,283,284,285]. Thus, several compounds of C. albidus may induce potential mechanisms resulting in an increased bioavailability. Furthermore, the absorption and bioavailability of C. albidus’ compounds can be influenced by other factors such as food matrix, processing (extraction), and individual variations.
8. Therapeutical Uses
8.1. Traditional Uses
Plant resources have always been an integral part of human society throughout history. Until the middle of the last century, traditional medicines provided an alternative and inexpensive source of primary health care for the rural population. However, with access to synthetic drugs, a large number of medicinal plants became obsolete, the memory of which in the population, after only two generations, is being lost.
One of these medicinal plants is C. albidus, which has been used in traditional folk medicine for a variety of illnesses [10,286,287,288], especially for the treatment of fever, diarrhea and other gastrointestinal illnesses [8], skin diseases, rheumatism, and various inflammatory diseases [182]. For the sake of completeness, it is mentioned here that C. albidus has also been used as a tanning agent [181], as an insect repellent, and as a substitute for tobacco, highly appreciated, moreover, for its hypotensive effect [49,288,289].
The decoction of leaves was traditionally used in the Spanish Levant as a tranquilizer, in the Baixa Plana as a sedative [10], and as a remedy against Parkinson’s symptoms in Mallorca [290,291]. To relieve toothache, mouthwashes were made with a decoction of its leaves and flowers. A sip of the resulting liquid, once cold, was kept in the mouth for some time [266,292,293,294]. In addition, the decoction of the aerial parts was used as an external antiseptic, for wounds and skin infections [293,294,295,296]. In the Spanish Basque Country, several uses were reported. For example, decoction was applied for the treatment of ulcers and for the treatment of gangrene, and fresh leaves were used directly on the wound for disinfection [297].
In the Mediterranean region, the decoction of the aerial parts (leaves, stems, and flowers) has been used to regulate blood pressure [298,299]. It has also been a frequent remedy for hemorrhoids and to treat bruises and varicose veins [290]. The decoction of flowers and leaves has also been popularly used as an analgesic for oral infections [293] and for hepatoprotection in Granada and Mallorca [290,298]. The decoction of the fresh aerial parts, including the flowers, was used as a remedy against colds and flu infections, and against bronchitis [9,290,299] and whooping cough [10]. In the Spanish peninsula, C. albidus decoction has also been used as a remedy for osteoarthritis in the province of Jaen [266] and for rheumatism in the Valencian community and the Province of Jaen [292,300]. In addition, it was used as an external antiseptic for wound healing and skin infections in the provinces of Castellon, Mallorca, and Almería [10,290,293], and in Morocco [263,265]. In Sardinia (Italy), a traditional use is reported in poultices and ointments, which were applied directly to the wound [301].
In cases of gastrointestinal infections, in Almería (Spain), an infusion of dried leaves was prepared to reduce abdominal pain [293,298]. Against colic, in Castilla-La Mancha and Murcia (Spain) an infusion of young and tender shoots was administered, but it was also supplied by oral ingestion of the powder of dry leaves for treatment [9]. The dried leaf powder also served as an antidiarrheal in Jaen [266].
8.2. Scientific Evidence Confirming Traditional Uses
8.2.1. Antimicrobial Activity
The antimicrobial activity of C. albidus is attributed to two main compound classes, terpenes and polyphenols. Studies have shown that both terpenes and polyphenols have potent antibacterial and antifungal activities [12,46,177,274]. However, there are few studies on the antimicrobial activity of C. albidus.
The terpenoid fraction of C. albidus extracts has been found to exhibit antimicrobial activity against S. aureus, Bacillus subtilis, Listeria monocytogenes, Klebsiella pneumoniae, and Candida albicans [46]. Gram-positive bacteria are more affected by terpenoids than gram-negative bacteria [46]. The inhibitory activity against B. subtilis, S. aureus, L. monocytogenes, K. pneumoniae, and C. albicans is seen at a minimum inhibitory concentration (MIC50) of 20 µ/mL, while E. faecalis, E. coli, and E. freudii are not affected [46].
The polyphenolic compounds, such as gallic acid and some glycosylated derivatives of myricetin and quercetin, are found in C. albidus extracts that exhibit strong antimicrobial activity against S. aureus [46,177]. However, the specific role of polyphenols in the antimicrobial capacity of C. albidus extracts against several bacteria, including S. aureus and E. coli, is not yet clear. Some studies suggest a positive correlation between higher content of polyphenols and higher antibacterial activity against both Gram-positive and Gram-negative bacteria [177]. The antimicrobial activity may be attributed to a synergistic effect of different compounds, including phenolic and terpenoid. The polar fraction of C. albidus extracts, including the butanol extract, the spray-dried aqueous extract, and the ethyl acetate extract, all exhibit strong antimicrobial activity against S. aureus with MIC50 of 2.5 mg/mL [12], 60.0 µg/mL [177], and 1.25 mg/mL [12], respectively. The hydroalcoholic extract of C. albidus also shows strong activity against E. coli with MIC50 of 233 µg/mL [177].
8.2.2. Anti-Inflammatory, Antinociceptive, Analgesic, and Sedative Activity
The anti-inflammatory, antinociceptive, analgesic, and sedative effects reported in the traditional use of this species are probably based on both terpenes and polyphenols and may be the result of a synergy of both groups of compounds, each with different effects on the central nervous system (CNS). Among single compounds of the terpenoid class present in C. albidus with a reported neurophysiological activity are the monoterpenes bicyclogermacrene, borneol, p-cymene, germacrene-D, linalool, myrcene, α-phellandrene, safranal, and thymol and the sesquiterpenes abscisic acid, ⍺-bisabolol, β-caryophyllene, caryophyllenol, guaiol, selin-11-en-4-α-ol, β-sesquiphellandrene, and α-zingiberene (Table 1).
A recent study with α-zingiberene, one of the principal compounds in C. albidus, showed a significant anti-inflammatory activity resulting in a reduction in angiogenesis, macrophage activation, and the activity of metalloproteinases [165]. In a mice model, α-zingiberene was found to reduce neuroinflammation through histone deacetylase 1 (HDAC1) inhibition. Neuropathic pain results from microglia-spinal overexpression of HDAC1, and α-zingiberene was found to be a promising HDAC1 inhibitor, with an IC50 of 2.3 ± 0.1 µM. Further, the administration of α-zingiberene reduced thermal hyperalgesia and mechanical allodynia [163]. Moreover, germacrene-D, another principal compound, was shown to have the potential for the development of pharmaceutical formulations with anti-inflammatory, antinociceptive, and analgesic activities in non-toxic concentrations [144,304].
Due to the polyphenolic composition, significant anti-inflammatory and antinociceptive activities were observed in C. albidus extracts, especially for several flavonols [13,305]. Anxiolytic and analgesic properties have also been observed for apigenin diglucoside [184], which selectively binds with high affinity to GABAA receptors. Similarly, kaempferol, myricetin, and quercetin derivatives are potentially responsible for anxiolytic, antinociceptive, and anti-inflammatory activity without exerting psycho-modulatory effects [13,184,306,307,308]. In addition, quercetin and kaempferol exhibited antidepressant effects. It has been demonstrated in several studies that both compounds act as monoamine oxidase inhibitors [238,309].
In a murine model, a flavonol-enriched C. albidus extract exhibited a substantial reduction in paw oedema and significantly inhibited nitrite generation without affecting the cell viability of lipopolysaccharide-stimulated murine peritoneal macrophages [13]. The same study observed a downregulation of the proinflammatory enzymes cyclooxygenase (COX-2) and inducible nitric oxide synthase (iNOS) in macrophages treated with the C. albidus extract, as well as a decrease in p38 mitogen-activated protein kinases (MAPK) phosphorylation. Furthermore, a high antinociceptive activity was observed at a concentration of 100 mg/kg and proved to be as efficient as acetylsalicylic acid at concentrations of 200 mg/kg. An HPLC-DAD-ESI-MS/MS analysis of the used chloroform extract revealed that kaempferol and quercetin derivatives were potentially responsible for such effects [13]. It is suggested that these compounds may act through a CNS-mediated analgesic mechanism. Both effects suggest that the polyphenolic composition of this species may act on the central and peripheral nervous system through inhibition of the mechanisms of pain, acting as a sedative [13,310].
9. Potential Pharmacological Applications of C. albidus and Their Mechanisms
The pharmacological activity of traditional preparations of C. albidus is mainly based on the antimicrobial, anti-inflammatory, antinociceptive, and analgesic effects of several of its compounds. Most of these activities have been confirmed by recent studies as described in previous sections, but taking into account the whole spectrum of bioactive substances included in C. albidus samples, new applications derived from the above-mentioned pharmacological activities can be suggested, opening the field for future research and uses. In this context, labdane-type diterpenes are promising compounds that have been shown to exert strong antiviral activity. For example, manoyl oxide showed a high capacity to suppress the dengue virus. In a fraction of labdanum diethyl ether extract at a concentration of 31.25 μg/mL from C. creticus, the proliferation of this virus was suppressed by 100% [311].
In the field of cancer research, evidence suggests that 13-epi-manoyl oxide, a characteristic labdane-type diterpene of C. albidus, may play a pivotal role in antitumor activity, inducing apoptosis in leukemic cell lines through multiple pathways [170,312,313,314]. Flavanols such as prodelphinidins and epigallocatechin-gallocatechin-catechin oligomers were also shown to exert a significant growth-inhibitory activity against human prostate cell lines by blocking cell cycle partly at the G1/G0 phase and activating caspase-3, as was shown for prodelphinidins extracted from C. albidus [14], synthetic prodelphinidins [315], and oligomeric proanthocyanidins from Vigna angularis [316]. The cytotoxic effect was suggested to be related with the presence of the pyrogallol moiety [14]. However, due to the composition of C. albidus, future applications could focus on the prevention of neurodegenerative diseases.
9.1. Prevention of Neurodegenerative Diseases
A potential field of application regarding phenolic compounds from C. albidus is the prevention of neurodegenerative diseases (NDDs). The potential application of the phenolic compounds of C. albidus for NDD prevention is suggested based on a combination of properties such as free radical scavenging and heavy metal chelation.
9.1.1. Free Radical Scavenging
Pathogenesis of NDD is influenced by oxidative stress [317]. Studies have shown the preventive effects of antioxidants in this regard, and in particular, an improvement in cognitive and neural functions was observed [318]. In addition, human CNS studies found that antioxidants can improve cerebral blood flow (CBF) [319,320]. This effect is particularly significant since it influences the maintenance of cognitive performance through adult neurogenesis [321] by improving vascularity [322]. C. albidus is rich in antioxidants of both terpenes and polyphenols—especially the phenolic content, with high amounts of flavonols, phenolic acids, flavanols, anthocyanins, and tannins. However, although phenolic compounds are the most abundant and potent antioxidants in C. albidus, there are also non-phenolic compounds, such as terpenes, that contribute to the overall antioxidant activity of this species.
Within polyphenols, the flavonols myricetin and quercetin, both a hydroxylated form of kaempferol, are capable of inducing the enzyme glutathione-S-transferase, which is involved in resistance to oxidative stress. Myricetin and quercetin and their glycosides, especially the rhamnosides, act directly as free radical scavengers [323]. They are capable of preventing DNA, protein, and membrane damage due to their aromatic hydroxyl groups [324]. In particular, quercetin acts as a potent radical scavenger based on three structural parts—an O-dihydroxy structure (catechol in ring B), a 2,3-double bond in combination with a 4-oxo group, and the additional presence of a 3- and 5-hydroxyl group [178].
Furthermore, it was demonstrated that flavanols such as epicatechins and phenolic acids such as gallic acid exert strong neuroprotective effects. They are able to block the neurotoxic effects caused by oxidized proteins. For example, some HIV proteins are known to cause neurotoxicity in humans through mechanisms that activate macrophages and glial cells, inducing the production of oxidative stress. Epicatechins have been shown to neutralize this effect [325]. In another study, prior to gamma radiation exposure, epicatechin treatment prevented liver and testicular damage due to oxidative stress produced by free radical formation as a result of radiation [326], suggesting that epicatechin may present protective effects in patients undergoing radiotherapy. It was further shown that epicatechins decrease the susceptibility of low-density lipoproteins to oxidation, thus preventing the onset of atherosclerosis [327]. In addition, neuroinflammation in a PD-induced mouse model was significantly reduced when animals were given gallic acid (100 mg/kg) by attenuating heme oxygenase-1 (a redox-regulated protein) and α-synuclein aggregation (an indicator of neurodegeneration of the CNS), suggesting that gallic acid is capable of inhibiting lipopolysaccharide-induced oxidative stress and protein conjugation [328].
In general, several studies have demonstrated a positive correlation between the total phenolic content of C. albidus and its antioxidant activity [12,20,30,182,259]. Bouyahya et al. [274] determined the total phenolic content for an ethanolic extract of C. albidus as high as 112.48 ± 1.78 mg gallic acid equivalents per gram extract. The antioxidant activity (2,2-diphenyl-1-picrylhydrazyl radical scavenging assay, DDPH) was determined as well, with high values between 27.26 and 142 mg Trolox equivalents per gram dry weight by Gonçalves et al. [182] and Lukas et al. [11], respectively. A study of C. albidus from Sardinia showed the ability to scavenge DPPH radicals from C. albidus extracts with EC50 values of 31.93 μg mL−1 for the butanol fraction, 22.23 μg mL−1 for the ethyl acetate fraction, and 33.24 μg mL−1 for water [12].
Regarding the antioxidant activity of terpenes in C. albidus, these compounds are present in smaller but not negligible quantities and may be responsible for a synergistic effect. They may improve symptoms caused by inflammation processes, inhibiting them in different steps of the inflammatory process, as demonstrated by a series of studies [130,306,329,330]. The terpenes present in C. albidus with potential activity on oxidative stress are α-bisabolol, α-pinene, α-zingiberene, β-caryophyllene, δ-limonene, myrcene, p-cymene, linalool, and humulene [331]. The mechanisms of action involved in the antioxidant activity are mainly the reduction of lipid peroxidation induced by H2O2, decreasing the formation of ROS and the release of NO. Furthermore, terpenes can increase catalase, superoxide dismutase, and peroxidase activities; reduce glutathione content; and restore the mitochondrial membrane [332]. On the other hand, α-pinene, limonene, and β-caryophyllene are GABA receptor agonists, decreasing the activities of acetylcholinesterase and lipoxygenase. They can arrest the cell cycle in the G2/M phase [332].
It has been claimed that α-bisabolol exhibits antioxidant activity in chemical and/or biological assays. Studies have indicated that α-bisabolol significantly inhibits luminol-amplified chemiluminescence at concentrations ranging from 7.7 to 31 μg/mL for Candida albicans and N-formyl-methionyl-leucyl-phenylalanine, respectively. A similar effect was observed in the SIN-1 and H2O2/HOCl systems, suggesting that α-bisabolol is a means of enhancing antioxidant capacity [333].
Limonene, one of the most abundant terpenes in nature, was found to exert antidepressant-like effects in mice caused by reduced nitrite levels in the hippocampus [74]. In addition, it has been shown that limonene reduced the inflammatory response and decreased the levels of inflammatory cytokines such as IL-1, IL-6, and TNF-α, which are related to depressive symptoms [334].
In relation to linalool, there are several in vitro and in vivo studies that show strong anti-inflammatory and antioxidant activity, which confers it with important neuroprotective activity [335,336]. Its mechanism of action consists in reducing the activation of NF-κB and preventing its nuclear translocation [337].
9.1.2. Heavy Metal Chelation
Another factor associated with ROS is the accumulation of heavy metals in the CNS, which can be treated with chelation. These metals are essential for biological functions in plants and animals, but their chemical properties can cause toxicity by interfering with homeostasis. They may bind to inappropriate protein sites, displacing natural binding sites and causing cellular malfunction. Heavy metals have been found to cause oxidative damage to biological macromolecules, primarily by binding to DNA and nuclear proteins. Exposure to heavy metals is related to proinflammatory cytokines resulting in neuronal damage through neuroinflammation and can lead to various disorders and diseases [338].
Numerous studies have demonstrated a connection between AD, PD, and dementias, and an exposure to heavy metals due to oxidative stress caused by the formation of free radicals [339,340]. Results from several studies even suggest associations between the levels of metals during pregnancy and autism spectrum disorder (ASD) in children [341,342,343]. In a recent study, arsenic, cadmium, copper, mercury, manganese, magnesium, and lead were identified in the development of ASD. The results of this study suggest that the impact of these metals may be related, in addition, with attention deficit hyperactivity disorder (ADHD), sharing neurochemical and neurodevelopmental pathways [344].
Chelation is a process in which a molecule, known as a chelating agent, forms a complex with a metal ion by surrounding it and holding it in a stable, water-soluble form. Chelation can occur through the formation of coordinate covalent bonds between the metal ion and the chelating agent [345]. The metal chelation potential of polyphenols is highly dependent on catechol moieties and combinations of hydroxyl and carbonyl groups [346]. Combinations of these groups and moieties define metal-binding sites. Chelating agents are compounds that bind to metal ions and prevent them from participating in reactions that can generate reactive oxygen species (ROS). Metal ions can catalyze the production of ROS by participating in reactions that generate highly reactive oxygen species. By binding to these metal ions, chelating agents can prevent them from participating in these reactions, reducing the amount of ROS that are produced. Several polyphenols present in C. albidus may exert both antioxidant and chelating properties, as was shown for isolated polyphenols [347,348], and can form stable five- or six-membered rings with oxophilic transition metal ions such as iron, copper, and zinc, which are abundant in the CNS [348,349].
Flavonoids such as kaempferol, quercetin, myricetin, and luteolin, as well as flavanols such as catechins have multiple oxygen-containing functional groups, such as hydroxyl (-OH) and carboxyl (-COOH) groups, which are capable of binding to metal ions and forming chelates [348]. The chelating effect of these compounds may also be enhanced by their ability to form hydrogen bonds with metal ions, which further stabilizes the complex [350].
There is evidence that the free-radical-scavenging and metal-chelating effects of aqueous extracts of C. albidus could contribute to the prevention of Fe2+-induced lipid peroxidation [182]. Infusions of C. albidus showed a high capacity to form complexes with Fe2+ of 66.63% at a concentration of 1.6 mg mL−1. This is especially noteworthy because Fe2+ chelation was suggested to be a more important mechanism than direct free radical scavenging for the prevention of lipid peroxidation [182].
10. Toxicity
Aside from the fact that this species has been used as an insect repellent and its dead leaves are allelopathic to other plant species, to date, no intoxication produced by C. albidus has been reported for mammals. Studies to evaluate the toxicity of C. albidus extracts have not yet been performed.
For the genus Cistus, in general, there is no evidence of toxicity. However, isolated cases of intoxication in bovines have been associated with a possible ingestion of C. salvifolius [351,352]. However, the responsible substances contained in some species of the genus Cistus, especially in C. salvifolius, which could cause toxic effects, are not yet clear [353] and may be related to their tannins. Tannins differ from other phenolic compounds by their ability to precipitate proteins, exerting a direct inhibition of digestive enzymes that leads to the formation of indigestible complexes for ruminants. Bacterial fermentation converts tannins into gallic acid and pyrogallol, both of which are nephrotoxic to cattle [351]. It is noteworthy that C. albidus was widely used in ethno-veterinary medicine for a variety of diseases [9,299,354]. For example, a decoction of leaves and flowers of C. albidus was popularly used to treat rumination syndrome in horses [9].
11. Conclusions
The composition of secondary metabolites of C. albidus, with more than 200 identified compounds, supports the widespread use of this species in folk medicine and indicates the therapeutic potential, with mainly anti-inflammatory, antimicrobial, antinociceptive, and analgesic properties.
The broad-spectrum antimicrobial effects of the C. albidus extracts summarized in this review support further exploration of the extracts and their components for novel approaches. Furthermore, its potential to regulate physiological processes in the nervous system makes this species an interesting candidate to be used for neurodegenerative disorders and diseases, especially in those diseases with an inflammatory or oxidative implication.
No references to metabolites that could exhibit toxicity have been found, nor are there known episodes of intoxication in humans. Based on ethnobotanical research, the paraphyletic varieties of the Cistus subgenus, especially C. creticus and C. albidus, were traditionally used for direct consumption of their leaves (dried and ground) or their extracts (as infusions and decoctions). Therefore, based also on its long traditional use, this plant could be considered safe.
In this work, it has been argued that the effectiveness of traditional preparations based on C. albidus may be based mainly on three factors. On the one hand, there is the pharmacological activity that derives from certain terpenes and polyphenols naturally present in C. albidus. On the other hand, increased bioavailability of phenolic and terpenoid compounds is suggested due to the presence of α-bisabolol, tocopherols, and fatty acids. Finally, since they are present in the same drug (aerial parts), all the compounds coexist in an equilibrium, which suggests a natural synergism. Hence, the entire drug should be more effective than the isolated substances that compose it. This natural composition of all the compounds found in this species makes Cistus albidus a medicinal plant with high potential, especially for treating and preventing neurodegenerative diseases and disorders.
We hope that this review will help lay the groundwork for further comprehensive pharmacological and pharmaceutical studies to better understand the clinical relevance and use of C. albidus and improve its legal status as a traditional medicinal plant.
Author Contributions
D.R.d.B. and E.B.-C. conceived the present idea and designed the review; D.R.d.B. wrote the manuscript with critical comments from E.B.-C. and A.R.-C.; methodology, D.R.d.B.; resources, D.R.d.B.; writing—original draft preparation, D.R.d.B.; writing—review and editing, E.B.-C., A.R.-C. and D.R.d.B.; supervision, E.B.-C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Spanish Ministry of Economy and Competitiveness (TED2021-129932B-C21 and PID2021-125188OB-C32) and Agencia Valenciana de la Innovación (INNEST/2022/103). E.B.-C. has been supported by the Requalification of the Spanish University System with a 2021/2023 grant.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Guzmán, B.; Vargas, P. Systematics, Character Evolution, and Biogeography of Cistus L. (Cistaceae) Based on ITS, TrnL-TrnF, and MatK Sequences. Mol. Phylogenet. Evol. 2005, 37, 644–660. [Google Scholar] [CrossRef] [PubMed]
- Schubert, R.; Wagner, G. Pflanzennamen Und Botanische Fachwörter: Botanisches Lexikon; 8., neubearbeitete und erw. Aufl.; Neumann-Neudamm: Melsungen, Germany, 1984; ISBN 978-3-7888-0421-3. [Google Scholar]
- Schmidt, W. Gehölze für Mediterrane Gärten; Hortus Mediterraneus; Ulmer: Stuttgart, Germany, 1999; ISBN 978-3-8001-6590-2. [Google Scholar]
- Coombes, A.J. Dictionary of Plant Names, New ed.; Hamlyn: London, UK, 1994; ISBN 978-0-600-58187-1. [Google Scholar]
- Barrajón-Catalán, E.; Fernández-Arroyo, S.; Roldán, C.; Guillén, E.; Saura, D.; Segura-Carretero, A.; Micol, V. A Systematic Study of the Polyphenolic Composition of Aqueous Extracts Deriving from Several Cistus Genus Species: Evolutionary Relationship: Polyphenolic Characterization of Cistus Aqueous Extracts. Phytochem. Anal. 2011, 22, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Ormeño, E.; Baldy, V.; Ballini, C.; Fernandez, C. Production and Diversity of Volatile Terpenes from Plants on Calcareous and Siliceous Soils: Effect of Soil Nutrients. J. Chem. Ecol. 2008, 34, 1219–1229. [Google Scholar] [CrossRef]
- Gülz, P.-G.; Herrmann, T.; Hangst, K. Leaf Trichomes in the Genus Cistus. Flora 1996, 191, 85–104. [Google Scholar] [CrossRef]
- Pardo de Santayana, M. Inventario Español de los Conocimientos Tradicionales Relativos de la Biodiversidad: Primera fase: Introducción, Metodología y Fichas; Ministerio para la Transición Ecológica: Madrid, Spain, 2014; ISBN 978-84-491-1401-4. [Google Scholar]
- Verde, A.; Fajardo, J.; Obón, C.; Cebrían, F.; Rivera, D. Guía de las Plantas Medicinales de Castilla-La Mancha; Altabán Ediciones: Albacete, Spain, 2008; ISBN 987-84-96465-53-4. [Google Scholar]
- Mulet Pascual, L. Estudio Etnobotánico de La Provincia de Castellón; Diputación de Castellón: Castellón, Spain, 1991; ISBN 978-84-86895-24-2. [Google Scholar]
- Lukas, B.; Jovanovic, D.; Schmiderer, C.; Kostas, S.; Kanellis, A.; Gómez Navarro, J.; Aytaç, Z.; Koç, A.; Sözen, E.; Novak, J. Intraspecific Genetic Diversity of Cistus creticus L. and Evolutionary Relationships to Cistus albidus L. (Cistaceae): Meeting of the Generations? Plants 2021, 10, 1619. [Google Scholar] [CrossRef]
- Mastino, P.; Marchetti, M.A.; Costa, J.; Juliano, C.; Usai, M. Analytical Profiling of Phenolic Compounds in Extracts of Three Cistus Species from Sardinia and Their Potential Antimicrobial and Antioxidant Activity. Chem. Biodivers. 2021, 18, e2100053. [Google Scholar] [CrossRef]
- Tahiri, O.; Atmani-Kilani, D.; Sanchez-Fidalgo, S.; Aparicio-Soto, M.; Alarcón-de-la-Lastra, C.; Barrajón-Catalán, E.; Micol, V.; Atmani, D. The Flavonol-Enriched Cistus albidus Chloroform Extract Possesses in Vivo Anti-Inflammatory and Anti-Nociceptive Activity. J. Ethnopharmacol. 2017, 209, 210–218. [Google Scholar] [CrossRef]
- Makabe, H.; Fujii, H.; Mori, M.; Matsumoto, K.; Ishihara, C.; Kawaguchi, K.; Kawahara, S.; Hattori, Y. Synthesis of Prodelphinidin Trimer Isolated from Cistus albidus and Its Growth Inhibitory Activity Against Human Prostate Cancer Cell Lines. Heterocycles 2016, 92, 1822. [Google Scholar] [CrossRef]
- Centenaro, G.; de Miguel, S.; Amouzgar, L.; Piñuela, Y.; Son, D.; Bonet, J.A.; de Aragón, J.M.; Dashevskaya, S.; Castaño, C.; Alday, J.G. Silvicultural Management and Altitude Prevail on Soil Properties and Fungal Community in Shaping Understorey Plant Communities in a Mediterranean Pine Forest. Sci. Total Environ. 2023, 858, 159860. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Jing, X.; Van Stan, J.T.; Plaza-Álvarez, P.A.; Gonzalez-Romero, J.; Peña, E.; Moya, D.; Antonio Zema, D.; de las Heras, J. Changes in Soil Functionality Eight Years after Fire and Post-Fire Hillslope Stabilisation in Mediterranean Forest Ecosystems. Geoderma 2022, 409, 115603. [Google Scholar] [CrossRef]
- El Mamoun, I.; Mouna, F.; Mohammed, A.; Najib, B.; Zine-El Abidine, T.; Abdelkarim, G.; Didier, B.; Laurent, L.; Abdelaziz, S. Zinc, Lead, and Cadmium Tolerance and Accumulation in Cistus libanotis, Cistus albidus and Cistus salviifolius: Perspectives on Phytoremediation. Remediat. J. 2020, 30, 73–80. [Google Scholar] [CrossRef]
- Civeyrel, L.; Leclercq, J.; Demoly, J.-P.; Agnan, Y.; Quèbre, N.; Pélissier, C.; Otto, T. Molecular Systematics, Character Evolution, and Pollen Morphology of Cistus and Halimium (Cistaceae). Plant Syst. Evol. 2011, 295, 23–54. [Google Scholar] [CrossRef]
- Brossa, R.; Pintó-Marijuan, M.; Francisco, R.; López-Carbonell, M.; Chaves, M.M.; Alegre, L. Redox Proteomics and Physiological Responses in Cistus albidus Shrubs Subjected to Long-Term Summer Drought Followed by Recovery. Planta 2015, 241, 803–822. [Google Scholar] [CrossRef] [PubMed]
- Casadesús, A.; Bouchikh, R.; Pérez-Llorca, M.; Munné-Bosch, S. Linking Jasmonates with Vitamin E Accumulation in Plants: A Case Study in the Mediterranean Shrub Cistus albidus L. Planta 2021, 253, 36. [Google Scholar] [CrossRef]
- Guzmán, B.; Lledó, M.D.; Vargas, P. Adaptive Radiation in Mediterranean Cistus (Cistaceae). PLoS ONE 2009, 4, e6362. [Google Scholar] [CrossRef] [PubMed]
- Hernández, I.; Miret, J.A.; Van Der Kelen, K.; Rombaut, D.; Van Breusegem, F.; Munné-Bosch, S. Zeatin Modulates Flower Bud Development and Tocopherol Levels in Cistus albidus (L.) Plants as They Age. Plant Biol. 2015, 17, 90–96. [Google Scholar] [CrossRef]
- Bertolasi, B.; Zago, L.; Gui, L.; Sitzia, T.; Vanetti, I.; Binelli, G.; Puppi, G.; Buldrini, F.; Pezzi, G. Phenological and Genetic Characterization of Mediterranean Plants at the Peripheral Range: The Case of Cistus albidus near Lake Garda. Flora 2019, 252, 26–35. [Google Scholar] [CrossRef]
- Grant, O.M.; McNeilly, T.; Incoll, L.D. Genetic Diversity of Cistus albidus in South-East Spain Does Not Relate to Mesoclimate. Funct. Plant Biol. 2006, 33, 247. [Google Scholar] [CrossRef]
- Guzmán, B.; Vargas, P. Long-Distance Colonization of the Western Mediterranean by Cistus ladanifer (Cistaceae) despite the Absence of Special Dispersal Mechanisms. J. Biogeogr. 2009, 36, 954–968. [Google Scholar] [CrossRef]
- Arrington, J.M.; Kubitzki, K. Cistaceae. In Flowering Plants Dicotyledons; Kubitzki, K., Bayer, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 62–70. ISBN 978-3-642-07680-0. [Google Scholar]
- Müller, M.; Siles, L.; Cela, J.; Munné-Bosch, S. Perennially Young: Seed Production and Quality in Controlled and Natural Populations of Cistus albidus Reveal Compensatory Mechanisms That Prevent Senescence in Terms of Seed Yield and Viability. J. Exp. Bot. 2014, 65, 287–297. [Google Scholar] [CrossRef]
- Pérez-Llorca, M.; Casadesús, A.; Müller, M.; Munné-Bosch, S. Leaf Orientation as Part of the Leaf Developmental Program in the Semi-Deciduous Shrub, Cistus albidus L.: Diurnal, Positional, and Photoprotective Effects During Winter. Front. Plant Sci. 2019, 10, 767. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Llorca, M.; Casadesús, A.; Munné-Bosch, S.; Müller, M. Contrasting Patterns of Hormonal and Photoprotective Isoprenoids in Response to Stress in Cistus albidus during a Mediterranean Winter. Planta 2019, 250, 1409–1422. [Google Scholar] [CrossRef] [PubMed]
- Munné-Bosch, S.; Jubany-Marí, T.; Alegre, L. Enhanced Photo- and Antioxidative Protection, and Hydrogen Peroxide Accumulation in Drought-Stressed Cistus clusii and Cistus albidus Plants. Tree Physiol. 2003, 23, 1–12. [Google Scholar] [CrossRef]
- Roy, J.; Sonie, L. Germination and Population Dynamics of Cistus Species in Relation to Fire. J. Appl. Ecol. 1992, 29, 647. [Google Scholar] [CrossRef]
- Casadesús, A.; Bouchikh, R.; Munné-Bosch, S. Contrasting Seasonal Abiotic Stress and Herbivory Incidence in Cistus albidus L. Plants Growing in Their Natural Habitat on a Mediterranean Mountain. J. Arid Environ. 2022, 206, 104842. [Google Scholar] [CrossRef]
- Cabezudo, B.; Pérez Latorre, A.V.; Navarro, T.; Nieto Caldera, J.M. Estudios Fenomorfológicos En La Vegetación Del Sur de España. II. Alcornocales Mesomediterráneos. (Montes de Málaga, Málaga). Acta Bot. Malacit. 1993, 18, 179–188. [Google Scholar] [CrossRef]
- Blasco, S.; Mateu, I. Flowering and Fruiting Phenology and Breeding System of Cistus albidus L. Acta Bot. Gallica 1995, 142, 245–251. [Google Scholar] [CrossRef]
- Siles, L.; Müller, M.; Cela, J.; Hernández, I.; Alegre, L.; Munné-Bosch, S. Marked Differences in Seed Dormancy in Two Populations of the Mediterranean Shrub, Cistus albidus L. Plant Ecol. Divers. 2017, 10, 231–240. [Google Scholar] [CrossRef]
- Rizzotto, M. Ricerche tassonomiche e corologiche sulle Cistaceae. 1: Il genere Cistus L. in Italia. Webbia 1979, 33, 343–378. [Google Scholar] [CrossRef]
- Robles, C.; Dutoit, T.; Bonin, G. Inhibition Mechanisms and Successional Processes: A Case Study of Cistus albidus L. in Provence. Ecosyst. Sustain. Dev. 1998, 1, 437–446. [Google Scholar]
- Thanos, A.C.; Geroghiou, K.; Kadis, C.; Pantazi, C. Cistaceae: A plant family with hard seeds. Isr. J. Bot. 1993, 41, 251–263. [Google Scholar]
- Trabaud, L.; Oustric, J. Heat Requirements for Seed Germination of Three Cistus Species in the Garrigue of Southern France. Flora 1989, 183, 321–325. [Google Scholar] [CrossRef]
- Trabaud, L.; Renard, P. Do light and litter influence the recruitment of cistus spp. Stands? Isr. J. Plant Sci. 1999, 47, 1–9. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. A Classification System for Seed Dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]
- Polunin, O.; Schauer, T.; Everard, B. Pflanzen Europas; BLV-Bestimmungsbuch; BLV(-Verl. Ges.): München, Germany, 1971; ISBN 978-3-405-10929-5. [Google Scholar]
- Robles, C.; Garzino, S. Essential Oil Composition of Cistus albidus Leaves. Phytochemistry 1998, 48, 1341–1345. [Google Scholar] [CrossRef]
- Castells, E.; Peñuelas, J. Is There a Feedback between N Availability in Siliceous and Calcareous Soils and Cistus albidus Leaf Chemical Composition? Oecologia 2003, 136, 183–192. [Google Scholar] [CrossRef]
- ISO Standard No. 9235:2013; International Organization for Standardization Aromatic Natural Raw Materials. ISO: Geneva, Switzerland, 2013.
- Bechlaghem, K.; Allali, H.; Benmehdi, H.; Aissaoui, N.; Flamini, G. Chemical Analysis of the Essential Oils of Three Cistus Species Growing in North- West of Algeria. Agric. Conspec. Sci. 2019, 84, 283–293. [Google Scholar]
- Llusià, J.; Peñuelas, J.; Ogaya, R.; Alessio, G. Annual and Seasonal Changes in Foliar Terpene Content and Emission Rates in Cistus albidus L. Submitted to Soil Drought in Prades Forest (Catalonia, NE Spain). Acta Physiol. Plant. 2010, 32, 387–394. [Google Scholar] [CrossRef]
- Palá-Paúl, J.; Velasco-Negueruela, A.; Pérez-Alonso, M.J.; Sanz, J. Seasonal Variation in Chemical Composition of Cistus albidus L. from Spain. J. Essent. Oil Res. 2005, 17, 19–22. [Google Scholar] [CrossRef]
- Maccioni, S.; Baldini, R.; Cioni, P.L.; Tebano, M.; Flamini, G. In Vivo Volatiles Emission and Essential Oils from Different Organs and Pollen Of Cistus albidus from Caprione (Eastern Liguria, Italy). Flavour Fragr. J. 2007, 22, 61–65. [Google Scholar] [CrossRef]
- Morales-Soto, A.; Oruna-Concha, M.J.; Elmore, J.S.; Barrajón-Catalán, E.; Micol, V.; Roldán, C.; Segura-Carretero, A. Volatile Profile of Spanish Cistus Plants as Sources of Antimicrobials for Industrial Applications. Ind. Crops Prod. 2015, 74, 425–433. [Google Scholar] [CrossRef]
- Ormeño, E.; Mévy, J.P.; Vila, B.; Bousquet-Mélou, A.; Greff, S.; Bonin, G.; Fernandez, C. Water Deficit Stress Induces Different Monoterpene and Sesquiterpene Emission Changes in Mediterranean Species. Relationship between Terpene Emissions and Plant Water Potential. Chemosphere 2007, 67, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Ormeño, E.; Bousquet-Mélou, A.; Mévy, J.-P.; Greff, S.; Robles, C.; Bonin, G.; Fernandez, C. Effect of Intraspecific Competition and Substrate Type on Terpene Emissions from Some Mediterranean Plant Species. J. Chem. Ecol. 2007, 33, 277–286. [Google Scholar] [CrossRef]
- Paolini, J.; Tomi, P.; Bernardini, A.-F.; Bradesi, P.; Casanova, J.; Kaloustian, J. Detailed Analysis of the Essential Oil from Cistus albidus L. by Combination of GC/RI, GC/MS and 13C-NMR Spectroscopy. Nat. Prod. Res. 2008, 22, 1270–1278. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Gao, X. Brain Targeted Drug Delivery Systems: A Focus on Nanotechnology and and Nanoparticulates; Academic Press: London, UK, 2019; ISBN 978-0-12-814002-4. [Google Scholar]
- Xu, H.; Blair, N.T.; Clapham, D.E. Camphor Activates and Strongly Desensitizes the Transient Receptor Potential Vanilloid Subtype 1 Channel in a Vanilloid-Independent Mechanism. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 8924–8937. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Vermaak, I.; Viljoen, A. Camphor—A Fumigant during the Black Death and a Coveted Fragrant Wood in Ancient Egypt and Babylon—A Review. Molecules 2013, 18, 5434–5454. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Welsch, C.W.; Rao, A.R. Modulatory Influence of Camphor on the Activities of Hepatic Carcinogen Metabolizing Enzymes and the Levels of Hepatic and Extrahepatic Reduced Glutathione in Mice. Cancer Lett. 1995, 88, 163–169. [Google Scholar] [CrossRef]
- Laude, E.A.; Morice, A.H.; Grattan, T.J. The Antitussive Effects of Menthol, Camphor and Cineole in Conscious Guinea-Pigs. Pulm. Pharmacol. 1994, 7, 179–184. [Google Scholar] [CrossRef]
- Liu, H.; Zhou, Y.; Sun, Y.; Sheng, X.; Zhang, J.; Zhang, Z.; Ding, J. Effect of Menthol and Camphor on Permeation of Compound Diphenhydramine Cream in Vitro. Cent South Pharm. 2011, 9, 97–101. [Google Scholar]
- Patra, J.K.; Das, G.; Bose, S.; Banerjee, S.; Vishnuprasad, C.N.; Pilar Rodriguez-Torres, M.; Shin, H. Star Anise (Illicium verum): Chemical Compounds, Antiviral Properties, and Clinical Relevance. Phytother. Res. 2020, 34, 1248–1267. [Google Scholar] [CrossRef]
- Jeong, J.-G.; Kim, Y.S.; Min, Y.K.; Kim, S.H. Low Concentration of 3-Carene Stimulates the Differentiation of Mouse Osteoblastic MC3T3-E1 Subclone 4 Cells. Phytother. Res. 2008, 22, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Gil, M.L.; Jimenez, J.; Ocete, M.A.; Zarzuelo, A.; Cabo, M.M. Comparative Study of Different Essential Oils of Bupleurum gibraltaricum Lamarck. Pharmazie 1989, 44, 284–287. [Google Scholar] [PubMed]
- Pina-Vaz, C.; Gonçalves Rodrigues, A.; Pinto, E.; Costa-de-Oliveira, S.; Tavares, C.; Salgueiro, L.; Cavaleiro, C.; Gonçalves, M.J.; Martinez-de-Oliveira, J. Antifungal Activity of Thymus Oils and Their Major Compounds. J. Eur. Acad. Dermatol. Venereol. 2004, 18, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Aeschbach, R.; Löliger, J.; Scott, B.C.; Murcia, A.; Butler, J.; Halliwell, B.; Aruoma, O.I. Antioxidant Actions of Thymol, Carvacrol, 6-Gingerol, Zingerone and Hydroxytyrosol. Food Chem. Toxicol. 1994, 32, 31–36. [Google Scholar] [CrossRef]
- Jayakumar, S.; Madankumar, A.; Asokkumar, S.; Raghunandhakumar, S.; Gokula dhas, K.; Kamaraj, S.; Divya, M.G.J.; Devaki, T. Potential Preventive Effect of Carvacrol against Diethylnitrosamine-Induced Hepatocellular Carcinoma in Rats. Mol. Cell. Biochem. 2012, 360, 51–60. [Google Scholar] [CrossRef]
- Hotta, M.; Nakata, R.; Katsukawa, M.; Hori, K.; Takahashi, S.; Inoue, H. Carvacrol, a Component of Thyme Oil, Activates PPAR alpha and Gamma and Suppresses COX-2 Expression. J. Lipid Res. 2010, 51, 132–139. [Google Scholar] [CrossRef]
- Cho, S.; Choi, Y.; Park, S.; Park, T. Carvacrol Prevents Diet-Induced Obesity by Modulating Gene Expressions Involved in Adipogenesis and Inflammation in Mice Fed with High-Fat Diet. J. Nutr. Biochem. 2012, 23, 192–201. [Google Scholar] [CrossRef]
- Decker, K.; Keppler, D. Galactosamine Induced Liver Injury. Prog. Liver Dis. 1972, 4, 183–199. [Google Scholar]
- Boskabady, M.H.; Jandaghi, P. Relaxant Effects of Carvacrol on Guinea Pig Tracheal Chains and Its Possible Mechanisms. Pharmazie 2003, 58, 661–663. [Google Scholar]
- Peixoto-Neves, D.; Silva-Alves, K.S.; Gomes, M.D.M.; Lima, F.C.; Lahlou, S.; Magalhães, P.J.C.; Ceccatto, V.M.; Coelho-de-Souza, A.N.; Leal-Cardoso, J.H. Vasorelaxant Effects of the Monoterpenic Phenol Isomers, Carvacrol and Thymol, on Rat Isolated Aorta. Fundam. Clin. Pharmacol. 2010, 24, 341–350. [Google Scholar] [CrossRef]
- Quintans-Júnior, L.; Moreira, J.C.F.; Pasquali, M.A.B.; Rabie, S.M.S.; Pires, A.S.; Schröder, R.; Rabelo, T.K.; Santos, J.P.A.; Lima, P.S.S.; Cavalcanti, S.C.H.; et al. Antinociceptive Activity and Redox Profile of the Monoterpenes (+)-Camphene, p -Cymene, and Geranyl Acetate in Experimental Models. ISRN Toxicol. 2013, 2013, 459530. [Google Scholar] [CrossRef] [PubMed]
- Arabloei Sani, M.; Yaghmaei, P.; Hajebrahimi, Z.; Hayati Roodbari, N. Therapeutic Effect of P-Cymene on Lipid Profile, Liver Enzyme, and Akt/Mtor Pathway in Streptozotocin-Induced Diabetes Mellitus in Wistar Rats. J. Obes. 2022, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sun, J. D-Limonene: Safety and Clinical Applications. Altern. Med. Rev. J. Clin. Ther. 2007, 12, 259–264. [Google Scholar]
- Lorigooini, Z.; Boroujeni, S.N.; Sayyadi-Shahraki, M.; Rahimi-Madiseh, M.; Bijad, E.; Amini-Khoei, H. Limonene through Attenuation of Neuroinflammation and Nitrite Level Exerts Antidepressant-Like Effect on Mouse Model of Maternal Separation Stress. Behav. Neurol. 2021, 2021, 8817309. [Google Scholar] [CrossRef]
- Pattnaik, S.; Subramanyam, V.R.; Bapaji, M.; Kole, C.R. Antibacterial and Antifungal Activity of Aromatic Constituents of Essential Oils. Microbios 1997, 89, 39–46. [Google Scholar]
- Linck, V.M.; da Silva, A.L.; Figueiró, M.; Caramão, E.B.; Moreno, P.R.H.; Elisabetsky, E. Effects of Inhaled Linalool in Anxiety, Social Interaction and Aggressive Behavior in Mice. Phytomedicine 2010, 17, 679–683. [Google Scholar] [CrossRef]
- Jana, S.; Patra, K.; Sarkar, S.; Jana, J.; Mukherjee, G.; Bhattacharjee, S.; Mandal, D.P. Antitumorigenic Potential of Linalool Is Accompanied by Modulation of Oxidative Stress: An in Vivo Study in Sarcoma-180 Solid Tumor Model. Nutr. Cancer 2014, 66, 835–848. [Google Scholar] [CrossRef]
- Sugawara, Y.; Hara, C.; Aoki, T.; Sugimoto, N.; Masujima, T. Odor Distinctiveness between Enantiomers of Linalool: Difference in Perception and Responses Elicited by Sensory Test and Forehead Surface Potential Wave Measurement. Chem. Senses 2000, 25, 77–84. [Google Scholar] [CrossRef]
- Peana, A.T.; D’Aquila, P.S.; Panin, F.; Serra, G.; Pippia, P.; Moretti, M.D.L. Anti-Inflammatory Activity of Linalool and Linalyl Acetate Constituents of Essential Oils. Phytomedicine 2002, 9, 721–726. [Google Scholar] [CrossRef]
- Jansen, C.; Shimoda, L.M.N.; Kawakami, J.K.; Ang, L.; Bacani, A.J.; Baker, J.D.; Badowski, C.; Speck, M.; Stokes, A.J.; Small-Howard, A.L.; et al. Myrcene and Terpene Regulation of TRPV1. Channels 2019, 13, 344–366. [Google Scholar] [CrossRef]
- Bomfim, L.M.; Menezes, L.R.A.; Rodrigues, A.C.B.C.; Dias, R.B.; Gurgel Rocha, C.A.; Soares, M.B.P.; Neto, A.F.S.; Nascimento, M.P.; Campos, A.F.; Silva, L.C.R.C.e.; et al. Antitumour Activity of the Microencapsulation of Annona vepretorum Essential Oil. Basic Clin. Pharmacol. Toxicol. 2016, 118, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Sayyah, M.; Nadjafnia, L.; Kamalinejad, M. Anticonvulsant Activity and Chemical Composition of Artemisia dracunculus L. Essential Oil. J. Ethnopharmacol. 2004, 94, 283–287. [Google Scholar] [CrossRef]
- Pulaj, B.; Mustafa, B.; Nelson, K.; Quave, C.L.; Hajdari, A. Chemical Composition and in Vitro Antibacterial Activity of Pistacia terebinthus Essential Oils Derived from Wild Populations in Kosovo. BMC Complement. Altern. Med. 2016, 16, 147. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Sun, H.; Chen, S.; Zeng, L.; Wang, T. Anti-Fungal Activity, Mechanism Studies on α-Phellandrene and Nonanal against Penicillium cyclopium. Bot. Stud. 2017, 58, 13. [Google Scholar] [CrossRef] [PubMed]
- Piccinelli, A.C.; Santos, J.A.; Konkiewitz, E.C.; Oesterreich, S.A.; Formagio, A.S.N.; Croda, J.; Ziff, E.B.; Kassuya, C.A.L. Antihyperalgesic and Antidepressive Actions of (R)-(+)-Limonene, α-Phellandrene, and Essential Oil from Schinus terebinthifolius Fruits in a Neuropathic Pain Model. Nutr. Neurosci. 2015, 18, 217–224. [Google Scholar] [CrossRef]
- Lima, D.F.; Brandão, M.S.; Moura, J.B.; Leitão, J.M.R.S.; Carvalho, F.A.A.; Miúra, L.M.C.V.; Leite, J.R.S.A.; Sousa, D.P.; Almeida, F.R.C. Antinociceptive Activity of the Monoterpene α-Phellandrene in Rodents: Possible Mechanisms of Action. J. Pharm. Pharmacol. 2012, 64, 283–292. [Google Scholar] [CrossRef]
- Lin, J.-J.; Lu, K.-W.; Ma, Y.-S.; Tang, N.-Y.; Wu, P.-P.; Wu, C.-C.; Lu, H.-F.; Lin, J.-G.; Chung, J.-G. Alpha-Phellandrene, a Natural Active Monoterpene, Influences a Murine WEHI-3 Leukemia Model in Vivo by Enhancing Macrophague Phagocytosis and Natural Killer Cell Activity. In Vivo 2014, 28, 583–588. [Google Scholar]
- Karthikeyan, R.; Kanimozhi, G.; Prasad, N.R.; Agilan, B.; Ganesan, M.; Srithar, G. Alpha Pinene Modulates UVA-Induced Oxidative Stress, DNA Damage and Apoptosis in Human Skin Epidermal Keratinocytes. Life Sci. 2018, 212, 150–158. [Google Scholar] [CrossRef]
- Hou, J.; Zhang, Y.; Zhu, Y.; Zhou, B.; Ren, C.; Liang, S.; Guo, Y. α-Pinene Induces Apoptotic Cell Death via Caspase Activation in Human Ovarian Cancer Cells. Med. Sci. Monit. 2019, 25, 6631–6638. [Google Scholar] [CrossRef]
- Rodrigues, K.A.d.F.; Amorim, L.V.; Dias, C.N.; Moraes, D.F.C.; Carneiro, S.M.P.; Carvalho, F.A.d.A. Syzygium Cumini (L.) Skeels Essential Oil and Its Major Constituent α-Pinene Exhibit Anti-Leishmania Activity through Immunomodulation in Vitro. J. Ethnopharmacol. 2015, 160, 32–40. [Google Scholar] [CrossRef]
- Pinheiro, M.A.; Magalhães, R.; Torres, D.; Cavalcante, R.; Mota, F.X.; Oliveira Coelho, E.A.; Moreira, H.; Lima, G.; da Costa Araújo, P.; Cardoso, J.L.; et al. Gastroprotective Effect of Alpha-Pinene and Its Correlation with Antiulcerogenic Activity of Essential Oils Obtained from Hyptis Species. Pharmacogn. Mag. 2015, 11, 123. [Google Scholar] [CrossRef] [PubMed]
- Matthews Jucá, D.; da Silva, M.; Palheta Junior, R.; de Lima, F.; Okoba, W.; Lahlou, S.; de Oliveira, R.; dos Santos, A.; Magalhães, P. The Essential Oil of Eucalyptus tereticornis and Its Constituents, α- and β-Pinene, Show Accelerative Properties on Rat Gastrointestinal Transit. Planta Med. 2011, 77, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Utegenova, G.; Pallister, K.; Kushnarenko, S.; Özek, G.; Özek, T.; Abidkulova, K.; Kirpotina, L.; Schepetkin, I.; Quinn, M.; Voyich, J. Chemical Composition and Antibacterial Activity of Essential Oils from Ferula L. Species against Methicillin-Resistant Staphylococcus aureus. Molecules 2018, 23, 1679. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wu, N.; Zu, Y.; Fu, Y. Comparative Anti-Infectious Bronchitis Virus (IBV) Activity of (-)-Pinene: Effect on Nucleocapsid (N) Protein. Molecules 2011, 16, 1044–1054. [Google Scholar] [CrossRef] [PubMed]
- Zamyad, M.; Abbasnejad, M.; Esmaeili-Mahani, S.; Mostafavi, A.; Sheibani, V. The Anticonvulsant Effects of Ducrosia anethifolia (Boiss) Essential Oil Are Produced by Its Main Component Alpha-Pinene in Rats. Arq. Neuropsiquiatr. 2019, 77, 106–114. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, C.; Zhang, Q.; Shan, Y.; Gu, W.; Wang, S. Design, Synthesis and Biological Evaluation of Novel β-Pinene-Based Thiazole Derivatives as Potential Anticancer Agents via Mitochondrial-Mediated Apoptosis Pathway. Bioorg. Chem. 2019, 84, 468–477. [Google Scholar] [CrossRef]
- Liao, S.; Shang, S.; Shen, M.; Rao, X.; Si, H.; Song, J.; Song, Z. One-Pot Synthesis and Antimicrobial Evaluation of Novel 3-Cyanopyridine Derivatives of (−)-β-Pinene. Bioorg. Med. Chem. Lett. 2016, 26, 1512–1515. [Google Scholar] [CrossRef]
- Felipe, C.F.B.; Albuquerque, A.M.S.; de Pontes, J.L.X.; de Melo, J.Í.V.; Rodrigues, T.C.M.L.; de Sousa, A.M.P.; Monteiro, Á.B.; Ribeiro, A.E.d.S.; Lopes, J.P.; de Menezes, I.R.A.; et al. Comparative Study of Alpha- and Beta-Pinene Effect on PTZ-Induced Convulsions in Mice. Fundam. Clin. Pharmacol. 2019, 33, 181–190. [Google Scholar] [CrossRef]
- Assimopoulou, A.N.; Sinakos, Z.; Papageorgiou, V.P. Radical Scavenging Activity of Crocus sativus L. Extract and Its Bioactive Constituents. Phytother. Res. 2005, 19, 997–1000. [Google Scholar] [CrossRef]
- Pintado, C.; de Miguel, A.; Acevedo, O.; Nozal, L.; Novella, J.L.; Rotger, R. Bactericidal Effect of Saffron (Crocus sativus L.) on Salmonella enterica during Storage. Food Control 2011, 22, 638–642. [Google Scholar] [CrossRef]
- Hosseinzadeh, H.; Talebzadeh, F. Anticonvulsant Evaluation of Safranal and Crocin from Crocus sativus in Mice. Fitoterapia 2005, 76, 722–724. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, H.; Noraei, N.B. Anxiolytic and Hypnotic Effect of Crocus sativus Aqueous Extract and Its Constituents, Crocin and Safranal, in Mice. Phytother. Res. 2009, 23, 768–774. [Google Scholar] [CrossRef] [PubMed]
- Kianbakht, S.; Mozaffari, K. Effects of Saffron and Its Active Constituents, Crocin and Safranal, on Prevention of Indomethacin Induced Gastric Ulcers in Diabetic and Nondiabetic Rats. JMPIR 2009, 8, 30–38. [Google Scholar]
- Rudbäck, J.; Bergström, M.A.; Börje, A.; Nilsson, U.; Karlberg, A.-T. α-Terpinene, an Antioxidant in Tea Tree Oil, Autoxidizes Rapidly to Skin Allergens on Air Exposure. Chem. Res. Toxicol. 2012, 25, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Baldissera, M.D.; Grando, T.H.; Souza, C.F.; Gressler, L.T.; Stefani, L.M.; da Silva, A.S.; Monteiro, S.G. In Vitro and in Vivo Action of Terpinen-4-Ol, γ-Terpinene, and α-Terpinene against Trypanosoma evansi. Exp. Parasitol. 2016, 162, 43–48. [Google Scholar] [CrossRef]
- Fadel, H.; Kebbi, S.; Chalchat, J.-C.; Figueredo, G.; Chalard, P.; Benayache, F.; Ghedadba, N.; Benayache, S. Identification of Volatile Components and Antioxidant Assessment of the Aerial Part Extracts from an Algerian Cistus albidus L. of the Aures Region. J. New Technol. Mater. 2020, 10, 38–46. [Google Scholar] [CrossRef]
- Khaleel, C.; Tabanca, N.; Buchbauer, G. α-Terpineol, a Natural Monoterpene: A Review of Its Biological Properties. Open Chem. 2018, 16, 349–361. [Google Scholar] [CrossRef]
- Couladis, M.; Chinou, I.B.; Tzakou, O.; Petrakis, P.V. Composition and Antimicrobial Activity of the Essential Oil Of Hypericum rumeliacum Subsp.Apollinis (Boiss. & Heldr.). Phytother. Res. 2003, 17, 152–154. [Google Scholar] [CrossRef]
- Souza, R.; Cardoso, M.; Menezes, C.; Silva, J.; De Sousa, D.; Batista, J. Gastroprotective Activity of α-Terpineol in Two Experimental Models of Gastric Ulcer in Rats. Daru J. Fac. Pharm. Tehran Univ. Med. Sci. 2011, 19, 277–281. [Google Scholar]
- Nagoor Meeran, M.F.; Javed, H.; Al Taee, H.; Azimullah, S.; Ojha, S.K. Pharmacological Properties and Molecular Mechanisms of Thymol: Prospects for Its Therapeutic Potential and Pharmaceutical Development. Front. Pharmacol. 2017, 8, 380. [Google Scholar] [CrossRef]
- Nagoor Meeran, M.F.; Jagadeesh, G.S.; Selvaraj, P. Thymol, a Dietary Monoterpene Phenol Abrogates Mitochondrial Dysfunction in β-Adrenergic Agonist Induced Myocardial Infarcted Rats by Inhibiting Oxidative Stress. Chem. Biol. Interact. 2016, 244, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, S.; Pari, L. Role of Thymol on Hyperglycemia and Hyperlipidemia in High Fat Diet-Induced Type 2 Diabetic C57BL/6J Mice. Eur. J. Pharmacol. 2015, 761, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Mendes, S.S.; Bomfim, R.R.; Jesus, H.C.R.; Alves, P.B.; Blank, A.F.; Estevam, C.S.; Antoniolli, A.R.; Thomazzi, S.M. Evaluation of the Analgesic and Anti-Inflammatory Effects of the Essential Oil of Lippia gracilis Leaves. J. Ethnopharmacol. 2010, 129, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Bukovská, A.; Cikoš, Š.; Juhás, Š.; Il’ková, G.; Rehák, P.; Koppel, J. Effects of a Combination of Thyme and Oregano Essential Oils on TNBS-Induced Colitis in Mice. Mediators Inflamm. 2007, 1–9. [Google Scholar] [CrossRef]
- Bhandari, S.S.; Kabra, M.P. To Evaluate Anti-Anxiety Activity of Thymol. J. Acute Dis. 2014, 3, 136–140. [Google Scholar] [CrossRef]
- López-Carbonell, M.; Gabasa, M.; Jáuregui, O. Enhanced Determination of Abscisic Acid (ABA) and Abscisic Acid Glucose Ester (ABA-GE) in Cistus albidus Plants by Liquid Chromatography-Mass Spectrometry in Tandem Mode. Plant Physiol. Biochem. 2009, 47, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Llorca, M.; Caselles, V.; Müller, M.; Munné-Bosch, S. The Threshold between Life and Death in Cistus albidus L. Seedlings: Mechanisms Underlying Drought Tolerance and Resilience. Tree Physiol. 2021, 41, 1861–1876. [Google Scholar] [CrossRef]
- Bassaganya-Riera, J.; Skoneczka, J.; Kingston, D.G.J.; Krishnan, A.; Misyak, S.A.; Guri, A.J.; Pereira, A.; Carter, A.B.; Minorsky, P.; Tumarkin, R.; et al. Mechanisms of Action and Medicinal Applications of Abscisic Acid. Curr. Med. Chem. 2010, 17, 467–478. [Google Scholar] [CrossRef]
- Mollashahi, M.; Abbasnejad, M.; Esmaeili-Mahani, S. Phytohormone Abscisic Acid Elicits Antinociceptive Effects in Rats through the Activation of Opioid and Peroxisome Proliferator-Activated Receptors β/δ. Eur. J. Pharmacol. 2018, 832, 75–80. [Google Scholar] [CrossRef]
- Mulyaningsih, S.; Sporer, F.; Zimmermann, S.; Reichling, J.; Wink, M. Synergistic Properties of the Terpenoids Aromadendrene and 1,8-Cineole from the Essential Oil of Eucalyptus globulus against Antibiotic-Susceptible and Antibiotic-Resistant Pathogens. Phytomedicine 2010, 17, 1061–1066. [Google Scholar] [CrossRef]
- Yeo, S.K.; Ali, A.Y.; Hayward, O.A.; Turnham, D.; Jackson, T.; Bowen, I.D.; Clarkson, R. β-Bisabolene, a Sesquiterpene from the Essential Oil Extract of Opoponax (Commiphora guidottii), Exhibits Cytotoxicity in Breast Cancer Cell Lines: Anti-tumour properties of β-bisabolene. Phytother. Res. 2016, 30, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Maurya, A.K.; Singh, M.; Dubey, V.; Srivastava, S.; Luqman, S.; Bawankule, D.U. α-(-)-Bisabolol Reduces pro-Inflammatory Cytokine Production and Ameliorates Skin Inflammation. Curr. Pharm. Biotechnol. 2014, 15, 173–181. [Google Scholar] [CrossRef] [PubMed]
- van Zyl, R.L.; Seatlholo, S.T.; van Vuuren, S.F.; Viljoen, A.M. The Biological Activities of 20 Nature Identical Essential Oil Constituents. J. Essent. Oil Res. 2006, 18, 129–133. [Google Scholar] [CrossRef]
- Seki, T.; Kokuryo, T.; Yokoyama, Y.; Suzuki, H.; Itatsu, K.; Nakagawa, A.; Mizutani, T.; Miyake, T.; Uno, M.; Yamauchi, K.; et al. Antitumor Effects of α-Bisabolol against Pancreatic Cancer. Cancer Sci. 2011, 102, 2199–2205. [Google Scholar] [CrossRef]
- Ho, C.-L.; Liao, P.-C.; Wang, E.I.-C.; Su, Y.-C. Composition and Antifungal Activities of the Leaf Essential Oil of Neolitsea parvigemma from Taiwan. Nat. Prod. Commun. 2011, 6, 1357–1360. [Google Scholar] [CrossRef]
- Mastino, P.M.; Marchetti, M.; Costa, J.; Usai, M. Comparison of Essential Oils from Cistus Species Growing in Sardinia. Nat. Prod. Res. 2017, 31, 299–307. [Google Scholar] [CrossRef]
- Takei, M.; Umeyama, A.; Arihara, S. T-Cadinol and Calamenene Induce Dendritic Cells from Human Monocytes and Drive Th1 Polarization. Eur. J. Pharmacol. 2006, 537, 190–199. [Google Scholar] [CrossRef]
- Dahham, S.; Tabana, Y.; Iqbal, M.; Ahamed, M.; Ezzat, M.; Majid, A.; Majid, A. The Anticancer, Antioxidant and Antimicrobial Properties of the Sesquiterpene β-Caryophyllene from the Essential Oil of Aquilaria crassna. Molecules 2015, 20, 11808–11829. [Google Scholar] [CrossRef]
- Ojha, S.; Javed, H.; Azimullah, S.; Haque, M.E. β-Caryophyllene, a Phytocannabinoid Attenuates Oxidative Stress, Neuroinflammation, Glial Activation, and Salvages Dopaminergic Neurons in a Rat Model of Parkinson Disease. Mol. Cell. Biochem. 2016, 418, 59–70. [Google Scholar] [CrossRef]
- Bahi, A.; Al Mansouri, S.; Al Memari, E.; Al Ameri, M.; Nurulain, S.M.; Ojha, S. β-Caryophyllene, a CB2 Receptor Agonist Produces Multiple Behavioral Changes Relevant to Anxiety and Depression in Mice. Physiol. Behav. 2014, 135, 119–124. [Google Scholar] [CrossRef]
- de Oliveira, C.C.; de Oliveira, C.V.; Grigoletto, J.; Ribeiro, L.R.; Funck, V.R.; Grauncke, A.C.B.; de Souza, T.L.; Souto, N.S.; Furian, A.F.; Menezes, I.R.A.; et al. Anticonvulsant Activity of β-Caryophyllene against Pentylenetetrazol-Induced Seizures. Epilepsy Behav. 2016, 56, 26–31. [Google Scholar] [CrossRef]
- Fidyt, K.; Fiedorowicz, A.; Strządała, L.; Szumny, A. β-Caryophyllene and β-Caryophyllene Oxide-Natural Compounds of Anticancer and Analgesic Properties. Cancer Med. 2016, 5, 3007–3017. [Google Scholar] [CrossRef]
- Türkez, H.; Çelik, K.; Toğar, B. Effects of Copaene, a Tricyclic Sesquiterpene, on Human Lymphocytes Cells in Vitro. Cytotechnology 2014, 66, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Turkez, H.; Togar, B.; Tatar, A. Tricyclic Sesquiterpene Copaene Prevents H2O2-Induced Neurotoxicity. J. Intercult. Ethnopharmacol. 2014, 3, 21. [Google Scholar] [CrossRef]
- Park, S.Y.; Park, S.J.; Park, N.J.; Joo, W.H.; Lee, S.-J.; Choi, Y.-W. α-Iso-Cubebene Exerts Neuroprotective Effects in Amyloid Beta Stimulated Microglia Activation. Neurosci. Lett. 2013, 555, 143–148. [Google Scholar] [CrossRef]
- Lee, S.K.; Kim, S.D.; Lee, H.Y.; Baek, S.-H.; Ko, M.J.; Son, B.G.; Park, S.; Choi, Y.W.; Bae, Y.-S. α-Iso-Cubebene, a Natural Compound Isolated from Schisandra chinensis Fruit, Has Therapeutic Benefit against Polymicrobial Sepsis. Biochem. Biophys. Res. Commun. 2012, 426, 226–231. [Google Scholar] [CrossRef]
- Baek, S.E.; Jang, E.J.; Choi, J.M.; Choi, Y.W.; Kim, C.D. α-Iso-Cubebene Attenuates Neointima Formation by Inhibiting HMGB1-Induced Monocyte to Macrophage Differentiation via Suppressing ROS Production. Int. Immunopharmacol. 2022, 111, 109121. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, G.; Almanza, G.R.; Cheng, Y.; Peng, J.; Hamann, M.; Duan, R.-D.; Åkesson, B. Antiproliferative Effects of Curcuphenol, a Sesquiterpene Phenol. Fitoterapia 2010, 81, 762–766. [Google Scholar] [CrossRef] [PubMed]
- Bai, Z.; Yao, C.; Zhu, J.; Xie, Y.; Ye, X.-Y.; Bai, R.; Xie, T. Anti-Tumor Drug Discovery Based on Natural Product β-Elemene: Anti-Tumor Mechanisms and Structural Modification. Molecules 2021, 26, 1499. [Google Scholar] [CrossRef]
- Asakura, K.; Matsuo, Y.; Oshima, T.; Kihara, T.; Minagawa, K.; Araki, Y.; Kagawa, K.; Kanemasa, T.; Ninomiya, M. ω-Agatoxin IVA-Sensitive Ca2+ Channel Blocker, α-Eudesmol, Protects against Brain Injury after Focal Ischemia in Rats. Eur. J. Pharmacol. 2000, 394, 57–65. [Google Scholar] [CrossRef]
- Moon, P.-D.; Han, N.-R.; Lee, J.S.; Kim, H.-Y.; Hong, S.; Kim, H.-J.; Yoo, M.-S.; Kim, H.-M.; Jeong, H.-J. β-Eudesmol Inhibits Thymic Stromal Lymphopoietin through Blockade of Caspase-1/NF-ΚB Signal Cascade in Allergic Rhinitis Murine Model. Chem. Biol. Interact. 2018, 294, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Narahara, C.; Saeheng, T.; Chaijaroenkul, W.; Dumre, S.P.; Na-Bangchang, K.; Karbwang, J. β-Eudesmol Induces the Expression of Apoptosis Pathway Proteins in Cholangiocarcinoma Cell Lines. J. Res. Med. Sci. Off. J. Isfahan Univ. Med. Sci. 2020, 25, 7. [Google Scholar] [CrossRef]
- Dhyani, P.; Sati, P.; Sharma, E.; Attri, D.C.; Bahukhandi, A.; Tynybekov, B.; Szopa, A.; Sharifi-Rad, J.; Calina, D.; Suleria, H.A.R.; et al. Sesquiterpenoid Lactones as Potential Anti-Cancer Agents: An Update on Molecular Mechanisms and Recent Studies. Cancer Cell Int. 2022, 22, 305. [Google Scholar] [CrossRef] [PubMed]
- Guedes, J.B.; do Nascimento, A.L.; Costa, W.K.; de Veras, B.O.; de Aguiar, J.C.R.d.O.F.; Navarro, D.M.d.A.F.; Napoleão, T.H.; da Silva, M.V.; de Oliveira, A.M.; Correia, M.T.d.S. Eugenia gracillima Essential Oil Has Pharmaceutical Applications in Pain and Inflammation without Toxic Effects in Mice. J. Ethnopharmacol. 2023, 303, 115941. [Google Scholar] [CrossRef] [PubMed]
- Casiglia, S.; Bruno, M.; Bramucci, M.; Quassinti, L.; Lupidi, G.; Fiorini, D.; Maggi, F. Kundmannia sicula (L.) DC: A Rich Source of Germacrene D. J. Essent. Oil Res. 2017, 29, 437–442. [Google Scholar] [CrossRef]
- Mendes De Lacerda Leite, G.; De Oliveira Barbosa, M.; Pereira Lopes, M.J.; De Araújo Delmondes, G.; Bezerra, D.S.; Araújo, I.M.; Carvalho De Alencar, C.D.; Melo Coutinho, H.D.; Peixoto, L.R.; Barbosa-Filho, J.M.; et al. Pharmacological and Toxicological Activities of α-Humulene and Its Isomers: A Systematic Review. Trends Food Sci. Technol. 2021, 115, 255–274. [Google Scholar] [CrossRef]
- Sabir, U.; Irfan, H.M.; Alamgeer Ullah, A.; Althobaiti, Y.S.; Alshehri, F.S.; Niazi, Z.R. Downregulation of Hepatic Fat Accumulation, Inflammation and Fibrosis by Nerolidol in Purpose Built Western-Diet-Induced Multiple-Hit Pathogenesis of NASH Animal Model. Biomed. Pharmacother. 2022, 150, 112956. [Google Scholar] [CrossRef]
- Dong, J.-R.; Chang, W.-W.; Chen, S.-M. Nerolidol Inhibits Proliferation of Leiomyoma Cells via Reactive Oxygen Species-Induced DNA Damage and Downregulation of the ATM/Akt Pathway. Phytochemistry 2021, 191, 112901. [Google Scholar] [CrossRef]
- Misra, B.B.; Dey, S. Evaluation of in Vivo Anti-Hyperglycemic and Antioxidant Potentials of α-Santalol and Sandalwood Oil. Phytomedicine 2013, 20, 409–416. [Google Scholar] [CrossRef]
- Garlet, Q.I.; Souza, C.F.; Rodrigues, P.; Descovi, S.N.; Martinez-Rodríguez, G.; Baldisserotto, B.; Heinzmann, B.M. GABAa Receptor Subunits Expression in Silver Catfish (Rhamdia Quelen) Brain and Its Modulation by Nectandra grandiflora Nees Essential Oil and Isolated Compounds. Behav. Brain Res. 2019, 376, 112178. [Google Scholar] [CrossRef]
- Tyagi, A.K.; Prasad, S.; Yuan, W.; Li, S.; Aggarwal, B.B. Identification of a Novel Compound (β-Sesquiphellandrene) from Turmeric (Curcuma longa) with Anticancer Potential: Comparison with Curcumin. Investig. New Drugs 2015, 33, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, J.; Yang, B.; Lv, G.-P.; Li, S.-P. Free Radical Scavenging Activity and Characterization of Sesquiterpenoids in Four Species of Curcuma Using a TLC Bioautography Assay and GC-MS Analysis. Molecules 2010, 15, 7547–7557. [Google Scholar] [CrossRef] [PubMed]
- Martins, R.M.G.; Xavier-Júnior, F.H.; Barros, M.R.; Menezes, T.M.; de Assis, C.R.D.; de Melo, A.C.G.R.; Veras, B.O.; Ferraz, V.P.; Filho, A.A.M.; Yogui, G.T.; et al. Impact on Cholinesterase-Inhibition and in Silico Investigations of Sesquiterpenoids from Amazonian Siparuna guianensis Aubl. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2021, 252, 119511. [Google Scholar] [CrossRef] [PubMed]
- Manjima, R.B.; Ramya, S.; Kavithaa, K.; Paulpandi, M.; Saranya, T.; Harysh Winster, S.B.; Balachandar, V.; Arul, N. Spathulenol Attenuates 6-Hydroxydopamine Induced Neurotoxicity in SH-SY5Y Neuroblastoma Cells. Gene Rep. 2021, 25, 101396. [Google Scholar] [CrossRef]
- do Nascimento, K.F.; Moreira, F.M.F.; Alencar Santos, J.; Kassuya, C.A.L.; Croda, J.H.R.; Cardoso, C.A.L.; Vieira, M.d.C.; Góis Ruiz, A.L.T.; Ann Foglio, M.; de Carvalho, J.E.; et al. Antioxidant, Anti-Inflammatory, Antiproliferative and Antimycobacterial Activities of the Essential Oil of Psidium guineense Sw. and Spathulenol. J. Ethnopharmacol. 2018, 210, 351–358. [Google Scholar] [CrossRef]
- de Matos Balsalobre, N.; dos Santos, E.; Mariano dos Santos, S.; Arena, A.C.; Konkiewitz, E.C.; Ziff, E.B.; Nazari Formagio, A.S.; Leite Kassuya, C.A. Potential Anti-Arthritic and Analgesic Properties of Essential Oil and Viridiflorol Obtained from Allophylus edulis Leaves in Mice. J. Ethnopharmacol. 2023, 301, 115785. [Google Scholar] [CrossRef]
- Akiel, M.A.; Alshehri, O.Y.; Aljihani, S.A.; Almuaysib, A.; Bader, A.; Al-Asmari, A.I.; Alamri, H.S.; Alrfaei, B.M.; Halwani, M.A. Viridiflorol Induces Anti-Neoplastic Effects on Breast, Lung, and Brain Cancer Cells through Apoptosis. Saudi J. Biol. Sci. 2022, 29, 816–821. [Google Scholar] [CrossRef]
- Trevizan, L.N.F.; Nascimento, K.F.d.; Santos, J.A.; Kassuya, C.A.L.; Cardoso, C.A.L.; Vieira, M.d.C.; Moreira, F.M.F.; Croda, J.; Formagio, A.S.N. Anti-Inflammatory, Antioxidant and Anti-Mycobacterium tuberculosis Activity of Viridiflorol: The Major Constituent of Allophylus edulis (A. St.-Hil., A. Juss. & Cambess.) Radlk. J. Ethnopharmacol. 2016, 192, 510–515. [Google Scholar] [CrossRef]
- Oon, S.F.; Nallappan, M.; Tee, T.T.; Shohaimi, S.; Kassim, N.K.; Sa’ariwijaya, M.S.F.; Cheah, Y.H. Xanthorrhizol: A Review of Its Pharmacological Activities and Anticancer Properties. Cancer Cell Int. 2015, 15, 100. [Google Scholar] [CrossRef]
- Mordukhova, E.A.; Kim, D.; Kim, W.-G.; Hwang, J.-K.; Pan, J.-G. The Food-Grade Antimicrobial Xanthorrhizol Targets the Enoyl-ACP Reductase (FabI) in Escherichia coli. Bioorg. Med. Chem. Lett. 2020, 30, 127651. [Google Scholar] [CrossRef]
- Oon, S.F.; Nallappan, M.; Kassim, N.K.; Shohaimi, S.; Sa’ariwijaya, M.S.F.; Tee, T.T.; Cheah, Y.H. Hypolipidemic Activities of Xanthorrhizol Purified from Centrifugal TLC. Biochem. Biophys. Res. Commun. 2016, 478, 1403–1408. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y.; Hwang, J.-K.; Chun, H.S. Xanthorrhizol Attenuates Dextran Sulfate Sodium-Induced Colitis via the Modulation of the Expression of Inflammatory Genes in Mice. Life Sci. 2011, 88, 864–870. [Google Scholar] [CrossRef]
- Borgonetti, V.; Governa, P.; Manetti, F.; Galeotti, N. Zingiberene, a Non-Zinc-Binding Class I HDAC Inhibitor: A Novel Strategy for the Management of Neuropathic Pain. Phytomedicine 2023, 111, 154670. [Google Scholar] [CrossRef]
- Bou, D.; Lago, J.; Figueiredo, C.; Matsuo, A.; Guadagnin, R.; Soares, M.; Sartorelli, P. Chemical Composition and Cytotoxicity Evaluation of Essential Oil from Leaves of Casearia sylvestris, Its Main Compound α-Zingiberene and Derivatives. Molecules 2013, 18, 9477–9487. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, B.A.; Silva, R.F.; de Moura, F.B.R.; Narduchi, C.T.; Deconte, S.R.; Sartorelli, P.; Tomiosso, T.C.; Lago, J.H.G.; Araújo, F. de A. α-Zingiberene, a Sesquiterpene from Essential Oil from Leaves of Casearia sylvestris, Suppresses Inflammatory Angiogenesis and Stimulates Collagen Deposition in Subcutaneous Implants in Mice. Nat. Prod. Res. 2022, 36, 5858–5862. [Google Scholar] [CrossRef] [PubMed]
- Ravera, S.; Esposito, A.; Degan, P.; Caicci, F.; Manni, L.; Liguori, A.; Bisio, A.; Iobbi, V.; Schito, A.; Traverso, C.E.; et al. The diterpene manool extracted from Salvia tingitana lowers free radical production in retinal rod outer segments by inhibiting the extramitochondrial F1 F0 ATP synthase. Cell Biochem. Funct. 2021, 39, 528–535. [Google Scholar] [CrossRef]
- Nicolella, H.D.; de Oliveira, P.F.; Munari, C.C.; Costa, G.F.D.; Moreira, M.R.; Veneziani, R.C.S.; Tavares, D.C. Differential Effect of Manool—A Diterpene from Salvia officinalis, on Genotoxicity Induced by Methyl Methanesulfonate in V79 and HepG2 Cells. Food Chem. Toxicol. 2014, 72, 8–12. [Google Scholar] [CrossRef]
- Monteiro, A.S.e.N.; Campos, D.R.; Albuquerque, A.A.S.; Evora, P.R.B.; Ferreira, L.G.; Celotto, A.C. Efeito Do Diterpeno Manool Sobre a Pressão Arterial e Reatividade Vascular Em Ratos Normotensos e Hipertensos. Arq. Bras. Cardiol. 2020, 115, 669–677. [Google Scholar] [CrossRef]
- Castro, C.H.; Pontes, C.N.R. Efeitos Cardiovasculares Do Diterpeno Manool Em Ratos Normotensos e Hipertensos. Arq. Bras. Cardiol. 2020, 115, 678–679. [Google Scholar] [CrossRef]
- Angelopoulou, D.; Demetzos, C.; Dimas, C.; Perdetzoglou, D.; Loukis, A. Essential Oils and Hexane Extracts from Leaves and Fruits of Cistus monspeliensis. Cytotoxic Activity of Ent-13-Epi-Manoyl Oxide and Its Isomers. Planta Med. 2001, 67, 168–171. [Google Scholar] [CrossRef]
- Kim, J.-H.; Na, H.-J.; Kim, C.-K.; Kim, J.-Y.; Ha, K.-S.; Lee, H.; Chung, H.-T.; Kwon, H.J.; Kwon, Y.-G.; Kim, Y.-M. The Non-Provitamin A Carotenoid, Lutein, Inhibits NF-KappaB-Dependent Gene Expression through Redox-Based Regulation of the Phosphatidylinositol 3-Kinase/PTEN/Akt and NF-KappaB-Inducing Kinase Pathways: Role of H(2)O(2) in NF-KappaB Activation. Free Radic. Biol. Med. 2008, 45, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.-L.; Zhao, Y.-N.; Shi, Z.-Z.; Cong, D.; Bai, Y.-S. Lutein Inhibits Cell Growth and Activates Apoptosis via the PI3K/AKT/MTOR Signaling Pathway in A549 Human Non-Small-Cell Lung Cancer Cells. J. Environ. Pathol. Toxicol. Oncol. Off. 2018, 37, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ding, Y.; Niu, Q.; Xu, S.; Pang, L.; Ma, R.; Jing, M.; Feng, G.; Tang, J.X.; Zhang, Q.; et al. Lutein Has a Protective Effect on Hepatotoxicity Induced by Arsenic via Nrf2 Signaling. BioMed Res. Int. 2015, 2015, 315205. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, B.; Li, Z.; Ji, X.; Huang, J.; Zhang, H.; Jiang, C. The Protective Role of Lutein on Isoproterenol-Induced Cardiac Failure Rat Model through Improving Cardiac Morphology, Antioxidant Status via Positively Regulating Nrf2/HO-1 Signalling Pathway. Pharm. Biol. 2019, 57, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Okada, T.; Nakai, M.; Maeda, H.; Hosokawa, M.; Sashima, T.; Miyashita, K. Suppressive Effect of Neoxanthin on the Differentiation of 3T3-L1 Adipose Cells. J. Oleo Sci. 2008, 57, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Teuscher, E. Biogene Arzneimittel: Lehrbuch der Pharmazeutischen Biologie; 2020; ISBN 978-3-8047-3607-8. Available online: https://www.amazon.de/Biogene-Arzneimittel-Lehrbuch-Pharmazeutischen-Biologie/dp/3804736076 (accessed on 10 August 2023).
- Tomás-Menor, L.; Morales-Soto, A.; Barrajón-Catalán, E.; Roldán-Segura, C.; Segura-Carretero, A.; Micol, V. Correlation between the Antibacterial Activity and the Composition of Extracts Derived from Various Spanish Cistus Species. Food Chem. Toxicol. 2013, 55, 313–322. [Google Scholar] [CrossRef]
- Sticher, O.; Heilmann, J.; Zündorf, I.; Hänsel, R.; Steinegger, E. Pharmakognosie—Phytopharmazie; 10., völlig neu Bearbeitete Auflage.; Wissenschaftliche Verlagsgesellschaft: Stuttgart, Germany, 2015; ISBN 978-3-8047-3144-8. [Google Scholar]
- Belitz, H.-D.; Grosch, W.; Schieberle, P. Lehrbuch der Lebensmittelchemie; Springer-Lehrbuch; Sechste, Vollständig Überarbeitete Auflage; Springer: Berlin/Heidelberg, Germany, 2008; ISBN 978-3-540-73201-3. [Google Scholar]
- Qa’dan, F.; Petereit, F.; Nahrstedt, A. Prodelphinidin Trimers and Characterization of a Proanthocyanidin Oligomer from Cistus albidus. Pharmazie 2003, 58, 416–419. [Google Scholar]
- Wiesner, J.V. Die Rohstoffe des Pflanzenreiches; Verlag W. Engelmann: Leipzig, Berlin, Germany, 1921. [Google Scholar]
- Gonçalves, S.; Gomes, D.; Costa, P.; Romano, A. The Phenolic Content and Antioxidant Activity of Infusions from Mediterranean Medicinal Plants. Ind. Crops Prod. 2013, 43, 465–471. [Google Scholar] [CrossRef]
- Wang, M.; Firrman, J.; Liu, L.; Yam, K. A Review on Flavonoid Apigenin: Dietary Intake, ADME, Antimicrobial Effects, and Interactions with Human Gut Microbiota. BioMed Res. Int. 2019, 2019, 7010467. [Google Scholar] [CrossRef]
- Salgueiro, J.B.; Ardenghi, P.; Dias, M.; Ferreira, M.B.; Izquierdo, I.; Medina, J.H. Anxiolytic Natural and Synthetic Flavonoid Ligands of the Central Benzodiazepine Receptor Have No Effect on Memory Tasks in Rats. Pharmacol. Biochem. Behav. 1997, 58, 887–891. [Google Scholar] [CrossRef]
- Tyszka-Czochara, M.; Konieczny, P.; Majka, M. Caffeic Acid Expands Anti-Tumor Effect of Metformin in Human Metastatic Cervical Carcinoma HTB-34 Cells: Implications of AMPK Activation and Impairment of Fatty Acids De Novo Biosynthesis. Int. J. Mol. Sci. 2017, 18, 462. [Google Scholar] [CrossRef] [PubMed]
- Magnani, C.; Isaac, V.L.B.; Correa, M.A.; Salgado, H.R.N. Caffeic Acid: A Review of Its Potential Use in Medications and Cosmetics. Anal Methods 2014, 6, 3203–3210. [Google Scholar] [CrossRef]
- Alam, M.; Ahmed, S.; Elasbali, A.M.; Adnan, M.; Alam, S.; Hassan, M.I.; Pasupuleti, V.R. Therapeutic Implications of Caffeic Acid in Cancer and Neurological Diseases. Front. Oncol. 2022, 12, 860508. [Google Scholar] [CrossRef]
- Lopes, R.; Costa, M.; Ferreira, M.; Gameiro, P.; Fernandes, S.; Catarino, C.; Santos-Silva, A.; Paiva-Martins, F. Caffeic Acid Phenolipids in the Protection of Cell Membranes from Oxidative Injuries. Interaction with the Membrane Phospholipid Bilayer. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183727. [Google Scholar] [CrossRef]
- Agunloye, O.M.; Oboh, G.; Ademiluyi, A.O.; Ademosun, A.O.; Akindahunsi, A.A.; Oyagbemi, A.A.; Omobowale, T.O.; Ajibade, T.O.; Adedapo, A.A. Cardio-Protective and Antioxidant Properties of Caffeic Acid and Chlorogenic Acid: Mechanistic Role of Angiotensin Converting Enzyme, Cholinesterase and Arginase Activities in Cyclosporine Induced Hypertensive Rats. Biomed. Pharmacother. Biomed. Pharmacother. 2019, 109, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-Y.; Hong, C.-O.; Lee, G.P.; Kim, C.-T.; Lee, K.-W. The Hepatoprotection of Caffeic Acid and Rosmarinic Acid, Major Compounds of Perilla frutescens, against t-BHP-Induced Oxidative Liver Damage. Food Chem. Toxicol. 2013, 55, 92–99. [Google Scholar] [CrossRef]
- Kulkarni, N.P.; Vaidya, B.; Narula, A.S.; Sharma, S.S. Neuroprotective Potential of Caffeic Acid Phenethyl Ester (CAPE) in CNS Disorders: Mechanistic and Therapeutic Insights. Curr. Neuropharmacol. 2021, 19, 1401–1415. [Google Scholar] [CrossRef]
- Khaled, M.; Koriem, M. Caftaric Acid: An Overview on Its Structure, Daily Consumption, Bioavailability and Pharmacological Effects. Biointerface Res. Appl. Chem. 2020, 10, 5616–5623. [Google Scholar] [CrossRef]
- Gezer, C. Potential Health Effects of the Popular Compound of Artichoke: Cynarin (CYN). Prog. Nutr. 2017, 19. [Google Scholar] [CrossRef]
- Xia, N.; Pautz, A.; Wollscheid, U.; Reifenberg, G.; Förstermann, U.; Li, H. Artichoke, Cynarin and Cyanidin Downregulate the Expression of Inducible Nitric Oxide Synthase in Human Coronary Smooth Muscle Cells. Molecules 2014, 19, 3654–3668. [Google Scholar] [CrossRef]
- Sebastiani, G.; Almeida-Toledano, L.; Serra-Delgado, M.; Navarro-Tapia, E.; Sailer, S.; Valverde, O.; Garcia-Algar, O.; Andreu-Fernández, V. Therapeutic Effects of Catechins in Less Common Neurological and Neurodegenerative Disorders. Nutrients 2021, 13, 2232. [Google Scholar] [CrossRef] [PubMed]
- Jakubek, P.; Rajić, J.; Kuczyńska, M.; Suliborska, K.; Heldt, M.; Dziedziul, K.; Vidaković, M.; Namieśnik, J.; Bartoszek, A. Beyond Antioxidant Activity: Redox Properties of Catechins May Affect Changes in the DNA Methylation Profile—The Example of SRXN1 Gene. Antioxidants 2023, 12, 754. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Caleare, A.; Hensel, A.; Mello, J.C.P.; Pinha, A.B.; Panizzon, G.P.; Lechtenberg, M.; Petereit, F.; Nakamura, C.V. Flavan-3-Ols and Proanthocyanidins from Limonium brasiliense Inhibit the Adhesion of Porphyromonas gingivalis to Epithelial Host Cells by Interaction with Gingipains. Fitoterapia 2017, 118, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.; Kim, N.; Shin, Y.; Kim, S.-Y.; Kim, Y.-J. Activity of Catechins and Their Applications. Biomed. Dermatol. 2020, 4, 8. [Google Scholar] [CrossRef]
- Zduńska, K.; Dana, A.; Kolodziejczak, A.; Rotsztejn, H. Antioxidant Properties of Ferulic Acid and Its Possible Application. Skin Pharmacol. Physiol. 2018, 31, 332–336. [Google Scholar] [CrossRef]
- Pinheiro, P.G.; Santiago, G.M.P.; da Silva, F.E.F.; de Araújo, A.C.J.; de Oliveira, C.R.T.; Freitas, P.R.; Rocha, J.E.; Neto, J.B.d.A.; da Silva, M.M.C.; Tintino, S.R.; et al. Ferulic Acid Derivatives Inhibiting Staphylococcus aureus TetK and MsrA Efflux Pumps. Biotechnol. Rep. 2022, 34, e00717. [Google Scholar] [CrossRef]
- Liu, Y.; Shi, L.; Qiu, W.; Shi, Y. Ferulic Acid Exhibits Anti-Inflammatory Effects by Inducing Autophagy and Blocking NLRP3 Inflammasome Activation. Mol. Cell. Toxicol. 2022, 18, 509–519. [Google Scholar] [CrossRef]
- Ren, Z.; Zhang, R.; Li, Y.; Li, Y.; Yang, Z.; Yang, H. Ferulic Acid Exerts Neuroprotective Effects against Cerebral Ischemia/Reperfusion-Induced Injury via Antioxidant and Anti-Apoptotic Mechanisms in Vitro and in Vivo. Int. J. Mol. Med. 2017, 40, 1444–1456. [Google Scholar] [CrossRef]
- Antonopoulou, I.; Sapountzaki, E.; Rova, U.; Christakopoulos, P. Ferulic Acid From Plant Biomass: A Phytochemical With Promising Antiviral Properties. Front. Nutr. 2021, 8, 777576. [Google Scholar] [CrossRef]
- Lee, C.-C.; Wang, C.-C.; Huang, H.-M.; Lin, C.-L.; Leu, S.-J.; Lee, Y.-L. Ferulic Acid Induces Th1 Responses by Modulating the Function of Dendritic Cells and Ameliorates Th2-Mediated Allergic Airway Inflammation in Mice. Evid.-Based Complement. Altern. Med. 2015, 2015, 678487. [Google Scholar] [CrossRef]
- Esmat, M.A.; Osman, A.; Hassan, R.E.; Hagag, S.A.; El-Maghraby, T.K. Hepatoprotective Effect of Ferulic Acid and/or Low Doses of γ-Irradiation against Cisplatin-Induced Liver Injury in Rats. Hum. Exp. Toxicol. 2022, 41, 9603271221136204. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Yu, H.; Guo, W.; Kong, Y.; Gu, L.; Li, Q.; Yang, S.; Zhang, Y.; Wang, Y. The Anticancer Effects of Ferulic Acid Is Associated with Induction of Cell Cycle Arrest and Autophagy in Cervical Cancer Cells. Cancer Cell Int. 2018, 18, 102. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-H.; Park, J.-K.; Kim, K.-M.; Lee, H.-J.; Kim, S. In Vitro and in Vivo Antithrombotic and Cytotoxicity Effects of Ferulic Acid. J. Biochem. Mol. Toxicol. 2018, 32, e22004. [Google Scholar] [CrossRef]
- Narasimhan, A.; Chinnaiyan, M.; Karundevi, B. Ferulic Acid Exerts Its Antidiabetic Effect by Modulating Insulin-Signalling Molecules in the Liver of High-Fat Diet and Fructose-Induced Type-2 Diabetic Adult Male Rat. Appl. Physiol. Nutr. Metab. 2015, 40, 769–781. [Google Scholar] [CrossRef]
- Kroes, B.H.; van den Berg, A.J.; Quarles van Ufford, H.C.; van Dijk, H.; Labadie, R.P. Anti-Inflammatory Activity of Gallic Acid. Planta Med. 1992, 58, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Ojeaburu, S.I.; Oriakhi, K. Hepatoprotective, Antioxidant and, Anti-Inflammatory Potentials of Gallic Acid in Carbon Tetrachloride-Induced Hepatic Damage in Wistar Rats. Toxicol. Rep. 2021, 8, 177–185. [Google Scholar] [CrossRef]
- Dludla, P.V.; Nkambule, B.B.; Jack, B.; Mkandla, Z.; Mutize, T.; Silvestri, S.; Orlando, P.; Tiano, L.; Louw, J.; Mazibuko-Mbeje, S.E. Inflammation and Oxidative Stress in an Obese State and the Protective Effects of Gallic Acid. Nutrients 2018, 11, 23. [Google Scholar] [CrossRef]
- Zhang, T.; Ma, L.; Wu, P.; Li, W.; Li, T.; Gu, R.; Dan, X.; Li, Z.; Fan, X.; Xiao, Z. Gallic Acid Has Anticancer Activity and Enhances the Anticancer Effects of Cisplatin in Non-small Cell Lung Cancer A549 Cells via the JAK/STAT3 Signaling Pathway. Oncol. Rep. 2019, 41, 1779–1788. [Google Scholar] [CrossRef]
- Li, Z.-J.; Liu, M.; Dawuti, G.; Dou, Q.; Ma, Y.; Liu, H.-G.; Aibai, S. Antifungal Activity of Gallic Acid In Vitro and In Vivo. Phytother. Res. 2017, 31, 1039–1045. [Google Scholar] [CrossRef]
- Ahn, M.-J.; Kim, C.Y.; Lee, J.S.; Kim, T.G.; Kim, S.H.; Lee, C.-K.; Lee, B.-B.; Shin, C.-G.; Huh, H.; Kim, J. Inhibition of HIV-1 Integrase by Galloyl Glucoses from Terminalia chebula and Flavonol Glycoside Gallates from Euphorbia pekinensis. Planta Med. 2002, 68, 457–459. [Google Scholar] [CrossRef]
- Khan, A.N.; Singh, R.; Bhattacharya, A.; Chakravarti, R.; Roy, S.; Ravichandiran, V.; Ghosh, D. A Short Review on Glucogallin and Its Pharmacological Activities. Mini-Rev. Med. Chem. 2022, 22, 2820–2830. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.; Lakshmanapermalsamy, P.; Illuri, R.; Bhosle, D.; Sangli, G.K.; Mundkinajeddu, D. In Vitro Evaluation of Antioxidant Potential of Isolated Compounds and Various Extracts of Peel of Punica Granatum L. Pharmacogn. Res. 2018, 10, 44–48. [Google Scholar]
- Habtemariam, S. α-Glucosidase Inhibitory Activity of Kaempferol-3-O-Rutinoside. Nat. Prod. Commun. 2011, 6, 1934578X1100600. [Google Scholar] [CrossRef]
- Silva dos Santos, J.; Gonçalves Cirino, J.P.; de Oliveira Carvalho, P.; Ortega, M.M. The Pharmacological Action of Kaempferol in Central Nervous System Diseases: A Review. Front. Pharmacol. 2021, 11, 565700. [Google Scholar] [CrossRef]
- Tian, C.; Liu, X.; Chang, Y.; Wang, R.; Lv, T.; Cui, C.; Liu, M. Investigation of the Anti-Inflammatory and Antioxidant Activities of Luteolin, Kaempferol, Apigenin and Quercetin. S. Afr. J. Bot. 2021, 137, 257–264. [Google Scholar] [CrossRef]
- Imran, M.; Saeed, F.; Hussain, G.; Imran, A.; Mehmood, Z.; Gondal, T.A.; El-Ghorab, A.; Ahmad, I.; Pezzani, R.; Arshad, M.U.; et al. Myricetin: A Comprehensive Review on Its Biological Potentials. Food Sci. Nutr. 2021, 9, 5854–5868. [Google Scholar] [CrossRef]
- Oliveira, H.; Wu, N.; Zhang, Q.; Wang, J.; Oliveira, J.; de Freitas, V.; Mateus, N.; He, J.; Fernandes, I. Bioavailability Studies and Anticancer Properties of Malvidin Based Anthocyanins, Pyranoanthocyanins and Non-Oxonium Derivatives. Food Funct. 2016, 7, 2462–2468. [Google Scholar] [CrossRef]
- Shih, P.-H.; Wu, C.-H.; Yeh, C.-T.; Yen, G.-C. Protective Effects of Anthocyanins against Amyloid β-Peptide-Induced Damage in Neuro-2A Cells. J. Agric. Food Chem. 2011, 59, 1683–1689. [Google Scholar] [CrossRef]
- Decendit, A.; Mamani-Matsuda, M.; Aumont, V.; Waffo-Teguo, P.; Moynet, D.; Boniface, K.; Richard, E.; Krisa, S.; Rambert, J.; Mérillon, J.-M.; et al. Malvidin-3-O-β Glucoside, Major Grape Anthocyanin, Inhibits Human Macrophage-Derived Inflammatory Mediators and Decreases Clinical Scores in Arthritic Rats. Biochem. Pharmacol. 2013, 86, 1461–1467. [Google Scholar] [CrossRef]
- Rossetto, M.; Vanzani, P.; Mattivi, F.; Lunelli, M.; Scarpa, M.; Rigo, A. Synergistic Antioxidant Effect of Catechin and Malvidin 3-Glucoside on Free Radical-Initiated Peroxidation of Linoleic Acid in Micelles. Arch. Biochem. Biophys. 2002, 408, 239–245. [Google Scholar] [CrossRef]
- Kim, M.; Yin, J.; Hwang, I.H.; Park, D.H.; Lee, E.K.; Kim, M.J.; Lee, M.W. Anti-Acne Vulgaris Effects of Pedunculagin from the Leaves of Quercus mongolica by Anti-Inflammatory Activity and 5α-Reductase Inhibition. Molecules. 2020, 25, 2154. [Google Scholar] [CrossRef]
- Chang, J.H.; Cho, J.H.; Kim, H.H.; Lee, K.P.; Lee, M.W.; Han, S.S.; Lee, D.I. Antitumor Activity of Pedunculagin, One of the Ellagitannin. Arch. Pharm. Res. 1995, 18, 396–401. [Google Scholar] [CrossRef]
- Silva Fernandes, A.; Hollanda Véras, J.; Silva, L.S.; Puga, S.C.; Luiz Cardoso Bailão, E.F.; de Oliveira, M.G.; Cardoso, C.G.; Carneiro, C.C.; Costa Santos, S.D.; Chen-Chen, L. Pedunculagin Isolated from Plinia cauliflora Seeds Exhibits Genotoxic, Antigenotoxic and Cytotoxic Effects in Bacteria and Human Lymphocytes. J. Toxicol. Environ. Health A 2022, 85, 353–363. [Google Scholar] [CrossRef]
- Mattioli, R.; Francioso, A.; Mosca, L.; Silva, P. Anthocyanins: A Comprehensive Review of Their Chemical Properties and Health Effects on Cardiovascular and Neurodegenerative Diseases. Molecules 2020, 25, 3809. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Du, B.; Li, J.; Yang, Y.; Zhu, F. An Insight into Anti-Inflammatory Activities and Inflammation Related Diseases of Anthocyanins: A Review of Both In Vivo and In Vitro Investigations. Int. J. Mol. Sci. 2021, 22, 11076. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Deng, Z.; Chen, F.; Zheng, L.; Pan, Y.; Xing, Q.; Tsao, R.; Li, H. Synergistic Antioxidant Effects of Petunidin and Lycopene in H9c2 Cells Submitted to Hydrogen Peroxide: Role of Akt/Nrf2 Pathway. J. Food Sci. 2020, 85, 1752–1763. [Google Scholar] [CrossRef]
- Wang, Q.-Q.; Gao, H.; Yuan, R.; Han, S.; Li, X.-X.; Tang, M.; Dong, B.; Li, J.-X.; Zhao, L.-C.; Feng, J.; et al. Procyanidin A2, a Polyphenolic Compound, Exerts Anti-Inflammatory and Anti-Oxidative Activity in Lipopolysaccharide-Stimulated RAW264.7 Cells. PLoS ONE 2020, 15, e0237017. [Google Scholar] [CrossRef]
- Nawrot-Hadzik, I.; Matkowski, A.; Hadzik, J.; Dobrowolska-Czopor, B.; Olchowy, C.; Dominiak, M.; Kubasiewicz-Ross, P. Proanthocyanidins and Flavan-3-Ols in the Prevention and Treatment of Periodontitis-Antibacterial Effects. Nutrients 2021, 13, 165. [Google Scholar] [CrossRef]
- Choy, Y.Y.; Fraga, M.; Mackenzie, G.G.; Waterhouse, A.L.; Cremonini, E.; Oteiza, P.I. The PI3K/Akt Pathway Is Involved in Procyanidin-Mediated Suppression of Human Colorectal Cancer Cell Growth. Mol. Carcinog. 2016, 55, 2196–2209. [Google Scholar] [CrossRef]
- Tian, Y.; Yang, C.; Yao, Q.; Qian, L.; Liu, J.; Xie, X.; Ma, W.; Nie, X.; Lai, B.; Xiao, L.; et al. Procyanidin B2 Activates PPARγ to Induce M2 Polarization in Mouse Macrophages. Front. Immunol. 2019, 10, 1895. [Google Scholar] [CrossRef]
- Jung, H.A.; Ali, M.Y.; Bhakta, H.K.; Min, B.-S.; Choi, J.S. Prunin Is a Highly Potent Flavonoid from Prunus davidiana Stems That Inhibits Protein Tyrosine Phosphatase 1B and Stimulates Glucose Uptake in Insulin-Resistant HepG2 Cells. Arch. Pharm. Res. 2017, 40, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Rozadi, N.; Oktavia, S.; Fauziah, F. Pharmacological Activities of Punicalagin: A Review. J. Drug Deliv. Ther. 2022, 12, 148–155. [Google Scholar] [CrossRef]
- Kim, J.K.; Park, S.U. Quercetin and Its Role in Biological Functions: An Updated Review. EXCLI J. 2018, 17, 856–863. [Google Scholar] [CrossRef]
- Chimenti, F.; Cottiglia, F.; Bonsignore, L.; Casu, L.; Casu, M.; Floris, C.; Secci, D.; Bolasco, A.; Chimenti, P.; Granese, A.; et al. Quercetin as the Active Principle of Hypericum hircinum Exerts a Selective Inhibitory Activity against MAO-A: Extraction, Biological Analysis, and Computational Study. J. Nat. Prod. 2006, 69, 945–949. [Google Scholar] [CrossRef]
- Nguyen, T.L.A.; Bhattacharya, D. Antimicrobial Activity of Quercetin: An Approach to Its Mechanistic Principle. Molecules 2022, 27, 2494. [Google Scholar] [CrossRef]
- Rocha, M.F.G.; Sales, J.A.; da Rocha, M.G.; Galdino, L.M.; de Aguiar, L.; Pereira-Neto, W. de A.; de Aguiar Cordeiro, R.; Castelo-Branco, D. de S.C.M.; Sidrim, J.J.C.; Brilhante, R.S.N. Antifungal Effects of the Flavonoids Kaempferol and Quercetin: A Possible Alternative for the Control of Fungal Biofilms. Biofouling 2019, 35, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Kempuraj, D.; Madhappan, B.; Christodoulou, S.; Boucher, W.; Cao, J.; Papadopoulou, N.; Cetrulo, C.L.; Theoharides, T.C. Flavonols Inhibit Proinflammatory Mediator Release, Intracellular Calcium Ion Levels and Protein Kinase C Theta Phosphorylation in Human Mast Cells. Br. J. Pharmacol. 2005, 145, 934–944. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, Q.; Ma, W.; Tian, F.; Shen, H.; Zhou, M. A Combination of Quercetin and Resveratrol Reduces Obesity in High-Fat Diet-Fed Rats by Modulation of Gut Microbiota. Food Funct. 2017, 8, 4644–4656. [Google Scholar] [CrossRef]
- Cinkilic, N.; Cetintas, S.K.; Zorlu, T.; Vatan, O.; Yilmaz, D.; Cavas, T.; Tunc, S.; Ozkan, L.; Bilaloglu, R. Radioprotection by Two Phenolic Compounds: Chlorogenic and Quinic Acid, on X-Ray Induced DNA Damage in Human Blood Lymphocytes in Vitro. Food Chem. Toxicol. 2013, 53, 359–363. [Google Scholar] [CrossRef]
- Hur, J.Y.; Soh, Y.; Kim, B.H.; Suk, K.; Sohn, N.W.; Kim, H.C.; Kwon, H.C.; Lee, K.R.; Kim, S.Y. Neuroprotective and Neurotrophic Effects of Quinic Acids from Aster scaber in PC12 Cells. Biol. Pharm. Bull. 2001, 24, 921–924. [Google Scholar] [CrossRef]
- Jang, S.-A.; Park, D.W.; Kwon, J.E.; Song, H.S.; Park, B.; Jeon, H.; Sohn, E.-H.; Koo, H.J.; Kang, S.C. Quinic Acid Inhibits Vascular Inflammation in TNF-α-Stimulated Vascular Smooth Muscle Cells. Biomed. Pharmacother. Biomed. Pharmacother. 2017, 96, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Zanello, P.R.; Koishi, A.C.; Rezende Júnior, C.d.O.; Oliveira, L.A.; Pereira, A.A.; de Almeida, M.V.; Duarte dos Santos, C.N.; Bordignon, J. Quinic Acid Derivatives Inhibit Dengue Virus Replication in Vitro. Virol. J. 2015, 12, 223. [Google Scholar] [CrossRef] [PubMed]
- Alcázar Magaña, A.; Kamimura, N.; Soumyanath, A.; Stevens, J.F.; Maier, C.S. Caffeoylquinic Acids: Chemistry, Biosynthesis, Occurrence, Analytical Challenges, and Bioactivity. Plant J. 2021, 107, 1299–1319. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Yu, T.; Han, S.; Chen, X.; Wang, K.; Gao, P.; Li, L. Caffeoylquinic Acids Protect against Alcoholic Liver Injury and Rare C10 Acetylenic Acids from Erigeron Breviscapus. Nat. Prod. Res. 2023, 1–6. [Google Scholar] [CrossRef]
- Gong, G.; Guan, Y.-Y.; Zhang, Z.-L.; Rahman, K.; Wang, S.-J.; Zhou, S.; Luan, X.; Zhang, H. Isorhamnetin: A Review of Pharmacological Effects. Biomed. Pharmacother. 2020, 128, 110301. [Google Scholar] [CrossRef]
- Jnawali, H.N.; Jeon, D.; Jeong, M.-C.; Lee, E.; Jin, B.; Ryoo, S.; Yoo, J.; Jung, I.D.; Lee, S.J.; Park, Y.-M.; et al. Antituberculosis Activity of a Naturally Occurring Flavonoid, Isorhamnetin. J. Nat. Prod. 2016, 79, 961–969. [Google Scholar] [CrossRef]
- Guan, H.; Luo, W.; Bao, B.; Cao, Y.; Cheng, F.; Yu, S.; Fan, Q.; Zhang, L.; Wu, Q.; Shan, M. A Comprehensive Review of Rosmarinic Acid: From Phytochemistry to Pharmacology and Its New Insight. Molecules 2022, 27, 3292. [Google Scholar] [CrossRef]
- Tang, L.; Xiang, H.; Sun, Y.; Qiu, L.; Chen, D.; Deng, C.; Chen, W. Monopalmityloxy Shikimic Acid: Enzymatic Synthesis and Anticoagulation Activity Evaluation. Appl. Biochem. Biotechnol. 2009, 158, 408–415. [Google Scholar] [CrossRef]
- Veach, D.; Hosking, H.; Thompson, K.; Santhakumar, A.B. Anti-Platelet and Anti-Thrombogenic Effects of Shikimic Acid in Sedentary Population. Food Funct. 2016, 7, 3609–3616. [Google Scholar] [CrossRef]
- Fabre, N.; Urizzi, P.; Souchard, J.P.; Fréchard, A.; Claparols, C.; Fourasté, I.; Moulis, C. An Antioxidant Sinapic Acid Ester Isolated from Iberis Amara. Fitoterapia 2000, 71, 425–428. [Google Scholar] [CrossRef]
- Legrum, W. Riechstoffe, Zwischen Gestank und Duft: Vorkommen, Eigenschaften und Anwendung von Riechstoffen und deren Gemischen; Studienbücher Chemie; 1. Aufl.; Vieweg + Teubner: Wiesbaden, Germany, 2011; ISBN 978-3-8348-1245-2. [Google Scholar]
- Fahlbusch, K.-G.; Hammerschmidt, F.-J.; Panten, J.; Pickenhagen, W.; Schatkowski, D.; Bauer, K.; Garbe, D.; Surburg, H. Flavors and Fragrances. In Ullmann’s Encyclopedia of Industrial Chemistry; Wiley-VCH Verlag GmbH & Co. KGaA, Ed.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2003; p. a11_141. ISBN 978-3-527-30673-2. [Google Scholar]
- Fenaroli, G.; Burdock, G.A. Fenaroli’s Handbook of Flavor Ingredients, 4th ed.; CRC Press: Boca Raton, FL, USA, 2002; ISBN 978-0-8493-0946-5. [Google Scholar]
- Oñate, M.; Munné-Bosch, S. Loss of Flower Bud Vigour in the Mediterranean Shrub, Cistus albidus L. at Advanced Developmental Stages. Plant Biol. 2010, 12, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Jubany-Mari, T.; Munne-Bosch, S.; Lopez-Carbonell, M.; Alegre, L. Hydrogen Peroxide Is Involved in the Acclimation of the Mediterranean Shrub, Cistus albidus L., to Summer Drought. J. Exp. Bot. 2008, 60, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Wang, H. Epimerisation of Catechins in Green Tea Infusions. Food Chem. 2000, 70, 337–344. [Google Scholar] [CrossRef]
- Wang, R.; Zhou, W.; Jiang, X. Reaction Kinetics of Degradation and Epimerization of Epigallocatechin Gallate (EGCG) in Aqueous System over a Wide Temperature Range. J. Agric. Food Chem. 2008, 56, 2694–2701. [Google Scholar] [CrossRef] [PubMed]
- Saklar, S.; Ertas, E.; Ozdemir, I.S.; Karadeniz, B. Effects of Different Brewing Conditions on Catechin Content and Sensory Acceptance in Turkish Green Tea Infusions. J. Food Sci. Technol. 2015, 52, 6639–6646. [Google Scholar] [CrossRef]
- Alami Merrouni, I.; Kharchoufa, L.; Bencheikh, N.; Elachouri, M. Ethnobotanical Profile of Medicinal Plants Used by People of North-Eastern Morocco: Cross-Cultural and Historical Approach (Part I). Ethnobot. Res. Appl. 2021, 21, 1–45. [Google Scholar]
- Las Plantas En La Cultura Popular de La Provincia de Albacete; Fajardo, J.; Verde, A.; Rivera, D.; Obón, C. (Eds.) Serie I—Estudios; Instituto de Estudios Albacetenses “Don Juan Manuel” de la Excma; Diputacioń de Albacete: Albacete, Spain, 2000; ISBN 978-84-95394-08-8. [Google Scholar]
- Ajjoun, M.; Kharchoufa, L.; Alami Merrouni, I.; Elachouri, M. Moroccan Medicinal Plants Traditionally Used for the Treatment of Skin Diseases: From Ethnobotany to Clinical Trials. J. Ethnopharmacol. 2022, 297, 115532. [Google Scholar] [CrossRef]
- Fernández Ocaña, A.M. Estudio Etnobotánico En El Parque Natural de Las Sierras de Cazorla, Segura y Las Villas; Investigación Química de Un Grupo de Especies Interesantes, Departamento de Biología Animal, Vegetal y Ecología, Facultad de Ciencias Experimentales, Universidad de Jaén: Jáen, Spain, 2000. [Google Scholar]
- Fiorini, D.; Molle, A.; Nabissi, M.; Santini, G.; Benelli, G.; Maggi, F. Valorizing Industrial Hemp (Cannabis sativa L.) by-Products: Cannabidiol Enrichment in the Inflorescence Essential Oil Optimizing Sample Pre-Treatment Prior to Distillation. Ind. Crops Prod. 2019, 128, 581–589. [Google Scholar] [CrossRef]
- Ross, S.A.; ElSohly, M.A. The Volatile Oil Composition of Fresh and Air-Dried Buds of Cannabis sativa. J. Nat. Prod. 1996, 59, 49–51. [Google Scholar] [CrossRef]
- Antony, A.; Farid, M. Effect of Temperatures on Polyphenols during Extraction. Appl. Sci. 2022, 12, 2107. [Google Scholar] [CrossRef]
- Al Juhaimi, F.; Özcan, M.M.; Uslu, N.; Ghafoor, K. The Effect of Drying Temperatures on Antioxidant Activity, Phenolic Compounds, Fatty Acid Composition and Tocopherol Contents in Citrus Seed and Oils. J. Food Sci. Technol. 2018, 55, 190–197. [Google Scholar] [CrossRef]
- Larrauri, J.A.; Sánchez-Moreno, C.; Saura-Calixto, F. Effect of Temperature on the Free Radical Scavenging Capacity of Extracts from Red and White Grape Pomace Peels. J. Agric. Food Chem. 1998, 46, 2694–2697. [Google Scholar] [CrossRef]
- Alara, O.R.; Abdurahman, N.H.; Ukaegbu, C.I. Extraction of Phenolic Compounds: A Review. Curr. Res. Food Sci. 2021, 4, 200–214. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.-W.; Lin, L.-G.; Ye, W.-C. Techniques for Extraction and Isolation of Natural Products: A Comprehensive Review. Chin. Med. 2018, 13, 20. [Google Scholar] [CrossRef] [PubMed]
- Bouyahya, A. Determination of Phenol Content and Antibacterial Activity of Five Medicinal Plants Ethanolic Extracts from North-West of Morocco. J. Plant Pathol. Microbiol. 2016, 7, 4. [Google Scholar] [CrossRef]
- Reverchon, E.; Senatore, F. Isolation of Rosemary Oil: Comparison between Hydrodistillation and Supercritical CO2 Extraction. Flavour Fragr. J. 1992, 7, 227–230. [Google Scholar] [CrossRef]
- He, X.; Yang, J.; Huang, Y.; Zhang, Y.; Wan, H.; Li, C. Green and Efficient Ultrasonic-Assisted Extraction of Bioactive Components from Salvia miltiorrhiza by Natural Deep Eutectic Solvents. Molecules 2019, 25, 140. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and Bioefficacy of Polyphenols in Humans. I. Review of 97 Bioavailability Studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Morand, C.; Manach, C.; Rémésy, C. Absorption and Metabolism of Polyphenols in the Gut and Impact on Health. Biomed. Pharmacother. 2002, 56, 276–282. [Google Scholar] [CrossRef]
- Thilakarathna, S.; Rupasinghe, H. Flavonoid Bioavailability and Attempts for Bioavailability Enhancement. Nutrients 2013, 5, 3367–3387. [Google Scholar] [CrossRef]
- Failla, M.L.; Chitchumronchokchai, C.; Ferruzzi, M.G.; Goltz, S.R.; Campbell, W.W. Unsaturated Fatty Acids Promote Bioaccessibility and Basolateral Secretion of Carotenoids and α-Tocopherol by Caco-2 Cells. Food Funct. 2014, 5, 1101–1112. [Google Scholar] [CrossRef] [PubMed]
- Ferri, P.; Angelino, D.; Gennari, L.; Benedetti, S.; Ambrogini, P.; Del Grande, P.; Ninfali, P. Enhancement of Flavonoid Ability to Cross the Blood–Brain Barrier of Rats by Co-Administration with α-Tocopherol. Food Funct. 2015, 6, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-Z.; Lu, Y.; Du, S.-Y.; Shang, K.-X.; Cai, C.-B. Influence of Borneol and Muscone on Geniposide Transport through MDCK and MDCK-MDR1 Cells as Blood–Brain Barrier in Vitro Model. Int. J. Pharm. 2013, 456, 73–79. [Google Scholar] [CrossRef]
- Yin, Y.; Cao, L.; Ge, H.; Duanmu, W.; Tan, L.; Yuan, J.; Tunan, C.; Li, F.; Hu, R.; Gao, F.; et al. L-Borneol Induces Transient Opening of the Blood–Brain Barrier and Enhances the Therapeutic Effect of Cisplatin. NeuroReport 2017, 28, 506–513. [Google Scholar] [CrossRef]
- Yu, B.; Ruan, M.; Dong, X.; Yu, Y.; Cheng, H. The Mechanism of the Opening of the Blood–Brain Barrier by Borneol: A Pharmacodynamics and Pharmacokinetics Combination Study. J. Ethnopharmacol. 2013, 150, 1096–1108. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, W.; Chen, L.; Ma, S.; Ping, L.; Yang, Z. Enhancement of Intestinal Absorption of Akebia Saponin D by Borneol and Probenecid in Situ and in Vitro. Environ. Toxicol. Pharmacol. 2010, 29, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Alcaraz Ariza, F.J.; Le Houérou, H.-N. Flora Básica de la Región de Murcia; Sociedad Cooperativa de Enseñanza Severo Ochoa: Murcia, Spain, 2002; ISBN 978-84-600-9378-7. [Google Scholar]
- Stübing, G.; Peris, J.B. Plantas Medicinales de la Comunidad Valenciana; Generalitat Valenciana, Conselleria de Medio Ambiete: Valencia, Spain, 1998; ISBN 978-84-482-1805-8. [Google Scholar]
- Segarra i Durà, E. Etnobotànica Farmacèutica de Gàtova; Universitat de València: València, Spain, 2008; ISBN 978-84-370-6976-0. [Google Scholar]
- Ledesma, J. Estudio Del Uso Tradicional de Las Plantas Silvestres En La Sierra de Montsant; Trabajo Fin de Carrera, Escuela de Ingeniería Forestal de Lleida: Lleida, Spain, 2004. [Google Scholar]
- Carrió Cabrer, M.E. Contribució a l’etnobotànica de Mallorca; La Biodiversitat Vegetal i La Seva Gestió En Una Illa Mediterrània, Universidad de Barcelona: Barcelona, Spain, 2013. [Google Scholar]
- Carrió, E.; Vallès, J. Ethnobotany of Medicinal Plants Used in Eastern Mallorca (Balearic Islands, Mediterranean Sea). J. Ethnopharmacol. 2012, 141, 1021–1040. [Google Scholar] [CrossRef] [PubMed]
- Belda, A.; Zaragozí, B.; Belda, I.; Martínez, J.; Seva, E. Traditional Knowledge of Medicinal Plants in the Serra de Mariola Natural Park, South-Eastern Spain. Afr. J. Tradit. Complement. Altern. Med. 2013, 10, 299–309. [Google Scholar] [CrossRef]
- Martínez-Lirola, M.J.; González-Tejero, M.R.; Molero-Mesa, J. Ethnobotanical Resources in the Province of Almeria, Spain: Campos de Nijar. Econ. Bot. 1996, 50, 40–56. [Google Scholar] [CrossRef]
- Torres-Montes, F. Nombres y Usos Tradicionales de Las Plantas Silvestres En Almería (Estudio Linguistico y Etnografico); Diputación de Almería e Instituto de Estudios Almerienses: Almería, Spain, 2004; ISBN 978-84-8108-313-2. [Google Scholar]
- Alarcόn, R.; Pardo-de-Santayana, M.; Priestley, C.; Morales, R.; Heinrich, M. Medicinal and Local Food Plants in the South of Alava (Basque Country, Spain). J. Ethnopharmacol. 2015, 176, 207–224. [Google Scholar] [CrossRef]
- Ortuño Moya, I.M. Etnobotánica de Los Villares y Valdepeñas de Jaén (Sur de La Península Ibérica); Universidad de Jaén—Departamento de Biología Animal, Vegetal y Ecología, Facultad de Ciencias Experimentales: Jaén, Spain, 2004. [Google Scholar]
- Goicoetxea Marcaida, A. Etnobotánica y Medicina Popular en el País Vasco; Pastor: Madrid, Spain, 2017; ISBN 978-84-95461-61-2. [Google Scholar]
- González-Tejero García, M.R. Investigaciones Etnobotánicas En La Provincia de Granada; Departamento de Biología Vegetal, Facultad de Farmacia, Universidad de Granada: Granada, Spain, 1989. [Google Scholar]
- Rivera, D.; Alcaraz, F.; Verde, A.; Fajardo, J.; Obón, C. Las Plantas en la Cultura Popular; SOMEHN (Sociedad Mediterránea de Historia Natural): Murcia, Spain, 2008; ISBN 978-84-931786-0-4. [Google Scholar]
- Guzmán, A. Aproximación a La Etnobotánica de La Provincia de Jaén; Departamento de Biología Vegetal, Universidad de Granada: Granada, Spain, 1997. [Google Scholar]
- Atzei, A.D. Le Piante Nella Tradizione Popolare Della Sardegna: Documentazione Sugli Usi Alimentari, Aromatizzanti, Profumieri, Artigianali, Cosmetici, Medicinali, Veterinari, Magici, Ornamentali, Rituali, Religiosi, Tintori, Antiparassitari e Vari, Delle Piante; C. Delfino: Sassari, Italy, 2003; ISBN 978-88-7138-298-2. [Google Scholar]
- Rivera, D.; Robleda, A.; Obón, C.; Cano, F. Introducción al Mundo de Las Plantas Medicinales En Murcia, 1st ed.; Ayuntamiento de Murcia: Murcia, Spain, 1994; ISBN 978-84-606-1922-2. [Google Scholar]
- Pellicer, J. Costumari Botànic: Recerques Etnobotàniques a Les Comarques Centrals Valencianes; Farga monogràfica, 2nd ed.; Edicions del Bullent: Picanya, Spain, 2000; ISBN 978-84-96187-13-9. [Google Scholar]
- Ascari, J.; de Oliveira, M.S.; Nunes, D.S.; Granato, D.; Scharf, D.R.; Simionatto, E.; Otuki, M.; Soley, B.; Heiden, G. Chemical Composition, Antioxidant and Anti-Inflammatory Activities of the Essential Oils from Male and Female Specimens of Baccharis punctulata (Asteraceae). J. Ethnopharmacol. 2019, 234, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Yatoo, M.; Gopalakrishnan, A.; Saxena, A.; Parray, O.R.; Tufani, N.A.; Chakraborty, S.; Tiwari, R.; Dhama, K.; Iqbal, H.M.N. Anti-Inflammatory Drugs and Herbs with Special Emphasis on Herbal Medicines for Countering Inflammatory Diseases and Disorders—A Review. Recent Pat. Inflamm. Allergy Drug Discov. 2018, 12, 39–58. [Google Scholar] [CrossRef]
- Gertsch, J.; Leonti, M.; Raduner, S.; Racz, I.; Chen, J.-Z.; Xie, X.-Q.; Altmann, K.-H.; Karsak, M.; Zimmer, A. Beta-Caryophyllene Is a Dietary Cannabinoid. Proc. Natl. Acad. Sci. USA 2008, 105, 9099–9104. [Google Scholar] [CrossRef] [PubMed]
- Viola, H.; Wasowski, C.; Levi de Stein, M.; Wolfman, C.; Silveira, R.; Dajas, F.; Medina, J.; Paladini, A. Apigenin, a Component of Matricaria recutita Flowers, Is a Central Benzodiazepine Receptors-Ligand with Anxiolytic Effects. Planta Med. 1995, 61, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Vissiennon, C.; Nieber, K.; Kelber, O.; Butterweck, V. Route of Administration Determines the Anxiolytic Activity of the Flavonols Kaempferol, Quercetin and Myricetin—Are They Prodrugs? J. Nutr. Biochem. 2012, 23, 733–740. [Google Scholar] [CrossRef]
- Sloley, B.D.; Urichuk, L.J.; Morley, P.; Durkin, J.; Shan, J.J.; Pang, P.K.; Coutts, R.T. Identification of Kaempferol as a Monoamine Oxidase Inhibitor and Potential Neuroprotectant in Extracts of Ginkgo biloba Leaves. J. Pharm. Pharmacol. 2000, 52, 451–459. [Google Scholar] [CrossRef]
- Coppin, J.P.; Xu, Y.; Chen, H.; Pan, M.-H.; Ho, C.-T.; Juliani, R.; Simon, J.E.; Wu, Q. Determination of Flavonoids by LC/MS and Anti-Inflammatory Activity in Moringa oleifera. J. Funct. Foods 2013, 5, 1892–1899. [Google Scholar] [CrossRef]
- Kuchta, K.; Tung, N.H.; Ohta, T.; Uto, T.; Raekiansyah, M.; Grötzinger, K.; Rausch, H.; Shoyama, Y.; Rauwald, H.W.; Morita, K. The Old Pharmaceutical Oleoresin Labdanum of Cistus creticus L. Exerts Pronounced in Vitro Anti-Dengue Virus Activity. J. Ethnopharmacol. 2020, 257, 112316. [Google Scholar] [CrossRef]
- Dimas, K.; Kokkinopoulos, D.; Demetzos, C.; Vaos, B.; Marselos, M.; Malamas, M.; Tzavaras, T. The Effect of Sclareol on Growth and Cell Cycle Progression of Human Leukemic Cell Lines. Leuk. Res. 1999, 23, 217–234. [Google Scholar] [CrossRef]
- Dimas, K.; Demetzos, C.; Marsellos, M.; Sotiriadou, R.; Malamas, M.; Kokkinopoulos, D. Cytotoxic Activity of Labdane Type Diterpenes against Human Leukemic Cell Lines in Vitro. Planta Med. 1998, 64, 208–211. [Google Scholar] [CrossRef]
- Dimas, K.; Demetzos, C.; Mitaku, S.; Vaos, B.; Marselos, M.; Tzavaras, T.; Kokkinopoulos, D. Cytotoxic Activity and Antiproliferative Effects of a New Semi-Synthetic Derivative of Ent-3 Beta-Hydroxy-13-Epi-Manoyl Oxide on Human Leukemic Cell Lines. Anticancer Res. 1999, 19, 4065–4072. [Google Scholar] [PubMed]
- Fujii, W.; Toda, K.; Matsumoto, K.; Kawaguchi, K.; Kawahara, S.; Hattori, Y.; Fujii, H.; Makabe, H. Syntheses of Prodelphinidin B1, B2, and B4 and Their Antitumor Activities against Human PC-3 Prostate Cancer Cell Lines. Tetrahedron Lett. 2013, 54, 7188–7192. [Google Scholar] [CrossRef]
- Kawahara, S.-I.; Ishihara, C.; Matsumoto, K.; Senga, S.; Kawaguchi, K.; Yamamoto, A.; Suwannachot, J.; Hamauzu, Y.; Makabe, H.; Fujii, H. Identification and Characterization of Oligomeric Proanthocyanidins with Significant Anti-Cancer Activity in Adzuki Beans (Vigna angularis). Heliyon 2019, 5, e02610. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Manach, C.; Morand, C.; Rémésy, C.; Jiménez, L. Dietary Polyphenols and the Prevention of Diseases. Crit. Rev. Food Sci. Nutr. 2005, 45, 287–306. [Google Scholar] [CrossRef]
- Joseph, J.A.; Shukitt-Hale, B.; Denisova, N.A.; Bielinski, D.; Martin, A.; McEwen, J.J.; Bickford, P.C. Reversals of Age-Related Declines in Neuronal Signal Transduction, Cognitive, and Motor Behavioral Deficits with Blueberry, Spinach, or Strawberry Dietary Supplementation. J. Neurosci. 1999, 19, 8114–8121. [Google Scholar] [CrossRef]
- Francis, S.T.; Head, K.; Morris, P.G.; Macdonald, I.A. The Effect of Flavanol-Rich Cocoa on the FMRI Response to a Cognitive Task in Healthy Young People. J. Cardiovasc. Pharmacol. 2006, 47 (Suppl. S2), S215–S220. [Google Scholar] [CrossRef]
- Sorond, F.A.; Lipsitz, L.A.; Hollenberg, N.K.; Fisher, N.D.L. Cerebral Blood Flow Response to Flavanol-Rich Cocoa in Healthy Elderly Humans. Neuropsychiatr. Dis. Treat. 2008, 4, 433–440. [Google Scholar]
- Gage, F.H. Adult Neurogenesis in Mammals. Science 2019, 364, 827–828. [Google Scholar] [CrossRef]
- Boldrini, M.; Fulmore, C.A.; Tartt, A.N.; Simeon, L.R.; Pavlova, I.; Poposka, V.; Rosoklija, G.B.; Stankov, A.; Arango, V.; Dwork, A.J.; et al. Human Hippocampal Neurogenesis Persists throughout Aging. Cell Stem Cell 2018, 22, 589–599. [Google Scholar] [CrossRef]
- Mahmoudi, H.; Aouadhi, C.; Kaddour, R.; Gruber, M.; Zargouni, H.; Zaouali, W.; Hamida, N.B.; Nasri, M.B.; Ouerghi, Z.; Hosni, K. Comparison of Antioxidant and Antimicrobial Activities of Two Cultivated Cistus Species from Tunisia. Biosci. J. 2016, 32, 226–237. [Google Scholar] [CrossRef]
- Limasset, B.; le Doucen, C.; Dore, J.C.; Ojasoo, T.; Damon, M.; Crastes de Paulet, A. Effects of Flavonoids on the Release of Reactive Oxygen Species by Stimulated Human Neutrophils. Multivariate Analysis of Structure-Activity Relationships (SAR). Biochem. Pharmacol. 1993, 46, 1257–1271. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Wu, L.-J.; Tashiro, S.-I.; Onodera, S.; Ikejima, T. Elevated Levels of DNA Repair Enzymes and Antioxidative Enzymes by (+)-Catechin in Murine Microglia Cells after Oxidative Stress. J. Asian Nat. Prod. Res. 2006, 8, 61–71. [Google Scholar] [CrossRef]
- Cheng, Z.; Zhang, Z.; Han, Y.; Wang, J.; Wang, Y.; Chen, X.; Shao, Y.; Cheng, Y.; Zhou, W.; Lu, X.; et al. A Review on Anti-Cancer Effect of Green Tea Catechins. J. Funct. Foods 2020, 74, 104172. [Google Scholar] [CrossRef]
- Lavanchy, D. Evolving Epidemiology of Hepatitis C Virus. Clin. Microbiol. Infect. 2011, 17, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-L.; Hsu, C.-C.; Huang, H.-J.; Chang, C.-J.; Sun, S.-H.; Lin, A.M.-Y. Gallic Acid Attenuated LPS-Induced Neuroinflammation: Protein Aggregation and Necroptosis. Mol. Neurobiol. 2020, 57, 96–104. [Google Scholar] [CrossRef]
- Benito, C.; Tolón, R.M.; Pazos, M.R.; Núñez, E.; Castillo, A.I.; Romero, J. Cannabinoid CB2 Receptors in Human Brain Inflammation. Br. J. Pharmacol. 2008, 153, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Walter, L.; Franklin, A.; Witting, A.; Wade, C.; Xie, Y.; Kunos, G.; Mackie, K.; Stella, N. Nonpsychotropic Cannabinoid Receptors Regulate Microglial Cell Migration. J. Neurosci. 2003, 23, 1398–1405. [Google Scholar] [CrossRef]
- Gonzalez-Burgos, E.; Gomez-Serranillos, M.P. Terpene Compounds in Nature: A Review of Their Potential Antioxidant Activity. Curr. Med. Chem. 2012, 19, 5319–5341. [Google Scholar] [CrossRef]
- Kim, T.; Song, B.; Cho, K.S.; Lee, I.-S. Therapeutic Potential of Volatile Terpenes and Terpenoids from Forests for Inflammatory Diseases. Int. J. Mol. Sci. 2020, 21, 2187. [Google Scholar] [CrossRef]
- Braga, P.C.; Dal Sasso, M.; Fonti, E.; Culici, M. Antioxidant Activity of Bisabolol: Inhibitory Effects on Chemiluminescence of Human Neutrophil Bursts and Cell-Free Systems. Pharmacology 2009, 83, 110–115. [Google Scholar] [CrossRef]
- d’Alessio, P.A.; Bisson, J.-F.; Béné, M.C. Anti-Stress Effects of d-Limonene and Its Metabolite Perillyl Alcohol. Rejuvenation Res. 2014, 17, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Batista, P.A.; de Paula Werner, M.F.; Oliveira, E.C.; Burgos, L.; Pereira, P.; da Silva Brum, L.F.; Story, G.M.; Santos, A.R.S. The Antinociceptive Effect of (-)-Linalool in Models of Chronic Inflammatory and Neuropathic Hypersensitivity in Mice. J. Pain 2010, 11, 1222–1229. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lv, O.; Zhou, F.; Li, Q.; Wu, Z.; Zheng, Y. Linalool Inhibits LPS-Induced Inflammation in BV2 Microglia Cells by Activating Nrf2. Neurochem. Res. 2015, 40, 1520–1525. [Google Scholar] [CrossRef] [PubMed]
- Del Prado-Audelo, M.L.; Cortés, H.; Caballero-Florán, I.H.; González-Torres, M.; Escutia-Guadarrama, L.; Bernal-Chávez, S.A.; Giraldo-Gomez, D.M.; Magaña, J.J.; Leyva-Gómez, G. Therapeutic Applications of Terpenes on Inflammatory Diseases. Front. Pharmacol. 2021, 12, 704197. [Google Scholar] [CrossRef]
- Cicero, C.E.; Mostile, G.; Vasta, R.; Rapisarda, V.; Signorelli, S.S.; Ferrante, M.; Zappia, M.; Nicoletti, A. Metals and Neurodegenerative Diseases. A Systematic Review. Environ. Res. 2017, 159, 82–94. [Google Scholar] [CrossRef]
- Bakulski, K.M.; Seo, Y.A.; Hickman, R.C.; Brandt, D.; Vadari, H.S.; Hu, H.; Park, S.K. Heavy Metals Exposure and Alzheimer’s Disease and Related Dementias. J. Alzheimers Dis. 2020, 76, 1215–1242. [Google Scholar] [CrossRef]
- Pyatha, S.; Kim, H.; Lee, D.; Kim, K. Association between Heavy Metal Exposure and Parkinson’s Disease: A Review of the Mechanisms Related to Oxidative Stress. Antioxidants 2022, 11, 2467. [Google Scholar] [CrossRef]
- Adams, J.B.; Baral, M.; Geis, E.; Mitchell, J.; Ingram, J.; Hensley, A.; Zappia, I.; Newmark, S.; Gehn, E.; Rubin, R.A.; et al. The Severity of Autism Is Associated with Toxic Metal Body Burden and Red Blood Cell Glutathione Levels. J. Toxicol. 2009, 2009, 532640. [Google Scholar] [CrossRef]
- Arora, M.; Reichenberg, A.; Willfors, C.; Austin, C.; Gennings, C.; Berggren, S.; Lichtenstein, P.; Anckarsäter, H.; Tammimies, K.; Bölte, S. Fetal and Postnatal Metal Dysregulation in Autism. Nat. Commun. 2017, 8, 15493. [Google Scholar] [CrossRef]
- Grove, J.; Ripke, S.; Als, T.D.; Mattheisen, M.; Walters, R.; Won, H.; Pallesen, J.; Agerbo, E.; Andreassen, O.A.; Anney, R.; et al. Common Risk Variants Identified in Autism Spectrum Disorder; Common Risk Variants Identified in Autism Spectrum Disorder. bioRXiv 2017. [Google Scholar] [CrossRef]
- Skogheim, T.S.; Weyde, K.V.F.; Engel, S.M.; Aase, H.; Surén, P.; Øie, M.G.; Biele, G.; Reichborn-Kjennerud, T.; Caspersen, I.H.; Hornig, M.; et al. Metal and Essential Element Concentrations during Pregnancy and Associations with Autism Spectrum Disorder and Attention-Deficit/Hyperactivity Disorder in Children. Environ. Int. 2021, 152, 106468. [Google Scholar] [CrossRef] [PubMed]
- Hider, R.C.; Liu, Z.D.; Khodr, H.H. Metal Chelation of Polyphenols. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2001; Volume 335, pp. 190–203. ISBN 978-0-12-182236-1. [Google Scholar]
- Fraga, C.G.; Galleano, M.; Verstraeten, S.V.; Oteiza, P.I. Basic Biochemical Mechanisms behind the Health Benefits of Polyphenols. Mol. Aspects Med. 2010, 31, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Lawson, M.; Drostinova, L.; Lauro, P.; Poprac, P.; Brezova, V.; Michalik, M.; Lukes, V.; Valko, M. Protective Role of Quercetin against Copper(II)-Induced Oxidative Stress: A Spectroscopic, Theoretical and DNA Damage Study. Food Chem. Toxicol. 2017, 110, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Lakey-Beitia, J.; Burillo, A.M.; La Penna, G.; Hegde, M.L.; Rao, K.S. Polyphenols as Potential Metal Chelation Compounds Against Alzheimer’s Disease. J. Alzheimers Dis. 2021, 82, S335–S357. [Google Scholar] [CrossRef]
- Savelieff, M.G.; DeToma, A.S.; Derrick, J.S.; Lim, M.H. The Ongoing Search for Small Molecules to Study Metal-Associated Amyloid-β Species in Alzheimer’s Disease. Acc. Chem. Res. 2014, 47, 2475–2482. [Google Scholar] [CrossRef]
- Fernandez, M.T.; Mira, M.L.; Florêncio, M.H.; Jennings, K.R. Iron and Copper Chelation by Flavonoids: An Electrospray Mass Spectrometry Study. J. Inorg. Biochem. 2002, 92, 105–111. [Google Scholar] [CrossRef]
- Mignacca, S.A.; Mucciarelli, M.; Colombino, E.; Biasibetti, E.; Muscia, S.; Amato, B.; Presti, V.D.M.L.; Vazzana, I.; Galbo, A.; Capucchio, M.T. Cistus salviifolius Toxicity in Cattle. Vet. Pathol. 2020, 57, 115–121. [Google Scholar] [CrossRef]
- Yeruham, I.; Orgad, U.; Avidar, Y.; Perl, S.; Liberboim, M.; Adler, H.; Shlosberg, A. A Urinary Retention Syndrome in Beef Cows Probably Caused by Ingestion of Cistus. Rev. Méd Vét 2002, 153, 627–632. [Google Scholar]
- Bruno-Soares, A.M.; Abreu, J.M.; Soler, F. Preliminary in Vitro Studies of Cistus salvifolius Leaves in Relation to Metabolic Disorders in Sheep. In Poisonous Plants and Related Toxins; Acamovic, T., Stewart, C.S., Pennycott, T.W., Eds.; CABI: Wallingford, UK, 2004; pp. 56–62. ISBN 978-0-85199-614-1. [Google Scholar]
- Chaachouay, N.; Azeroual, A.; Douira, A.; Zidane, L. Ethnoveterinary Practices of Medicinal Plants among the Zemmour and Zayane Tribes, Middle Atlas, Morocco. S. Afr. J. Bot. 2022, 151, 826–840. [Google Scholar] [CrossRef]
Figure 1.
Typical Mediterranean landscape with a predominance of C. albidus, called jaral (left). Detail of flowers and leaves of C. albidus in spring (right). Images were captured by the authors.
Figure 1.
Typical Mediterranean landscape with a predominance of C. albidus, called jaral (left). Detail of flowers and leaves of C. albidus in spring (right). Images were captured by the authors.

Figure 2.
Trichomes: Glandular trichomes with secretion of bioactive compounds on their tops (left). Stellar trichomes covering the central vein of a leave (right). Images were captured by the authors using a Swift microscope, model SW380T-SC500-5PBC (M TEC USA Inc., Tschertz, TX, USA).
Figure 2.
Trichomes: Glandular trichomes with secretion of bioactive compounds on their tops (left). Stellar trichomes covering the central vein of a leave (right). Images were captured by the authors using a Swift microscope, model SW380T-SC500-5PBC (M TEC USA Inc., Tschertz, TX, USA).

Figure 3.
Natural distribution of Cistus albidus in the western Mediterranean basin (distribution map compiled using Google Earth (https://www.google.com/earth/download/, accessed on 19 April 2022). Highlighted in pink, the known natural distribution of C. albidus.
Figure 3.
Natural distribution of Cistus albidus in the western Mediterranean basin (distribution map compiled using Google Earth (https://www.google.com/earth/download/, accessed on 19 April 2022). Highlighted in pink, the known natural distribution of C. albidus.

Figure 4.
Systematics of the pink-purple flowered species of Cistus genus based on Guzmán and Vargas [1] and Civeyrel et al. [23].

Figure 5.
Structures of hydroxybenzoic and hydroxycinnamic acids.

Figure 6.
Basic structure of flavonoids with their three characteristic rings which are formed by different biosynthetic pathways. The B ring is derived from the shikimate pathway via phenylalanine.
Figure 6.
Basic structure of flavonoids with their three characteristic rings which are formed by different biosynthetic pathways. The B ring is derived from the shikimate pathway via phenylalanine.

Figure 7.
Structure of flavonoids with their three characteristic rings.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Raus de Baviera, D.; Ruiz-Canales, A.; Barrajón-Catalán, E. Cistus albidus L.—Review of a Traditional Mediterranean Medicinal Plant with Pharmacological Potential. Plants 2023, 12, 2988. https://doi.org/10.3390/plants12162988
AMA Style
Raus de Baviera D, Ruiz-Canales A, Barrajón-Catalán E. Cistus albidus L.—Review of a Traditional Mediterranean Medicinal Plant with Pharmacological Potential. Plants. 2023; 12(16):2988. https://doi.org/10.3390/plants12162988
Chicago/Turabian StyleRaus de Baviera, Daniel, Antonio Ruiz-Canales, and Enrique Barrajón-Catalán. 2023. "Cistus albidus L.—Review of a Traditional Mediterranean Medicinal Plant with Pharmacological Potential" Plants 12, no. 16: 2988. https://doi.org/10.3390/plants12162988
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.