Impact of Goji Berries (Lycium barbarum) Supplementation on the Energy Homeostasis of Rabbit Does: Uni- and Multivariate Approach

 ,
,  ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Body Conditions
2.3. Hormone and Metabolite Assays
2.4. Statistical Considerations and Data Analysis
3. Results
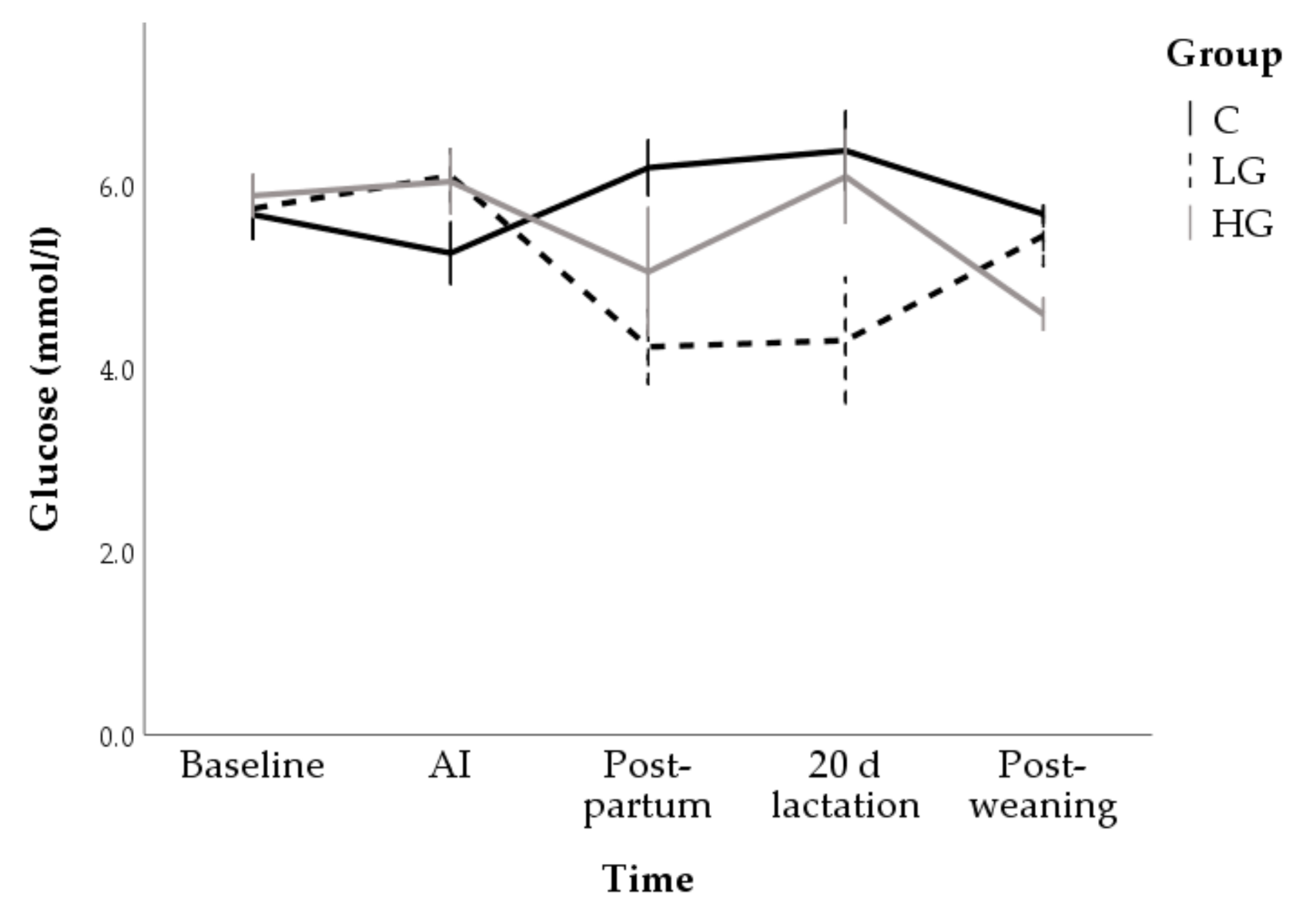
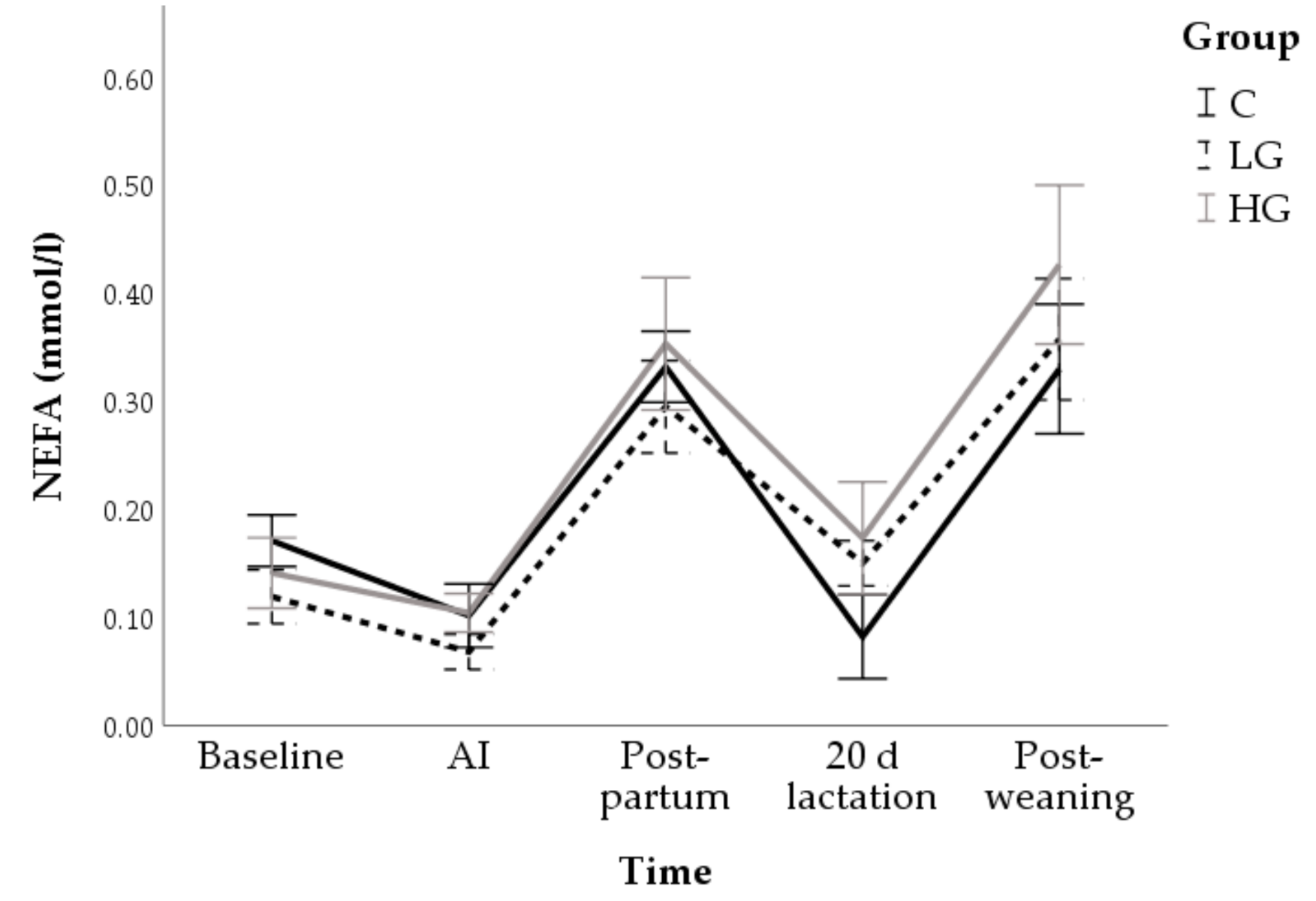
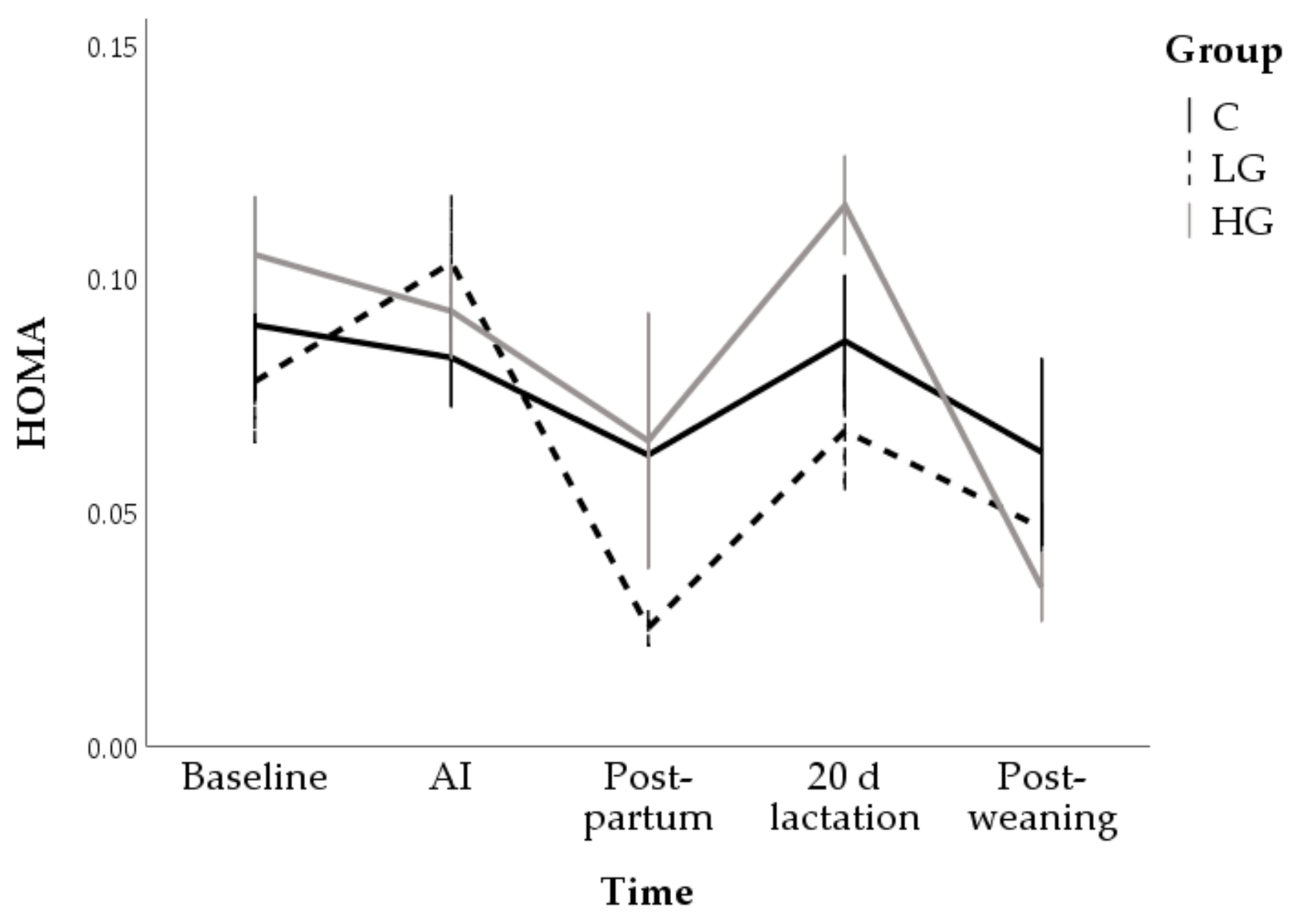
3.1. Univariable Approach
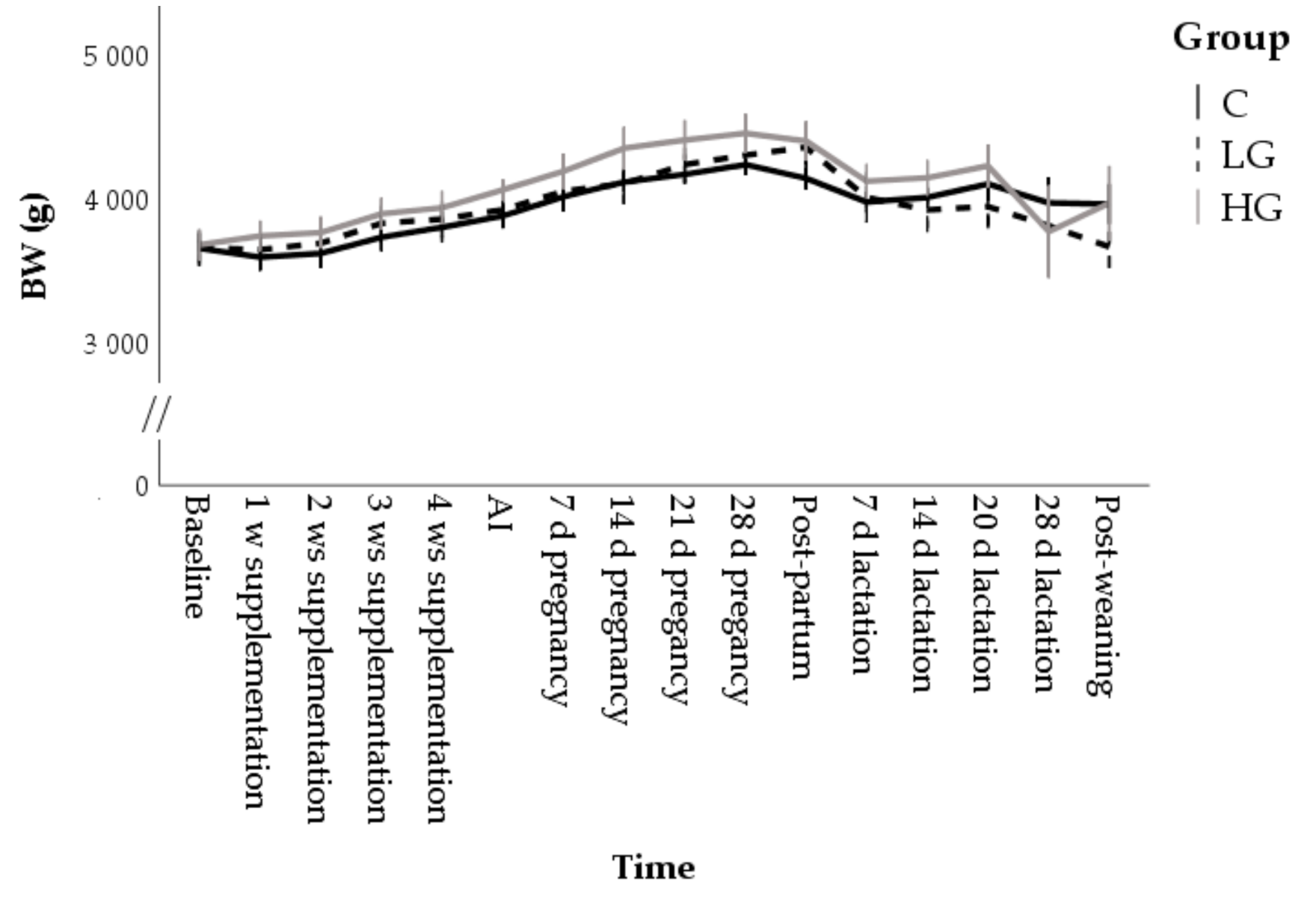
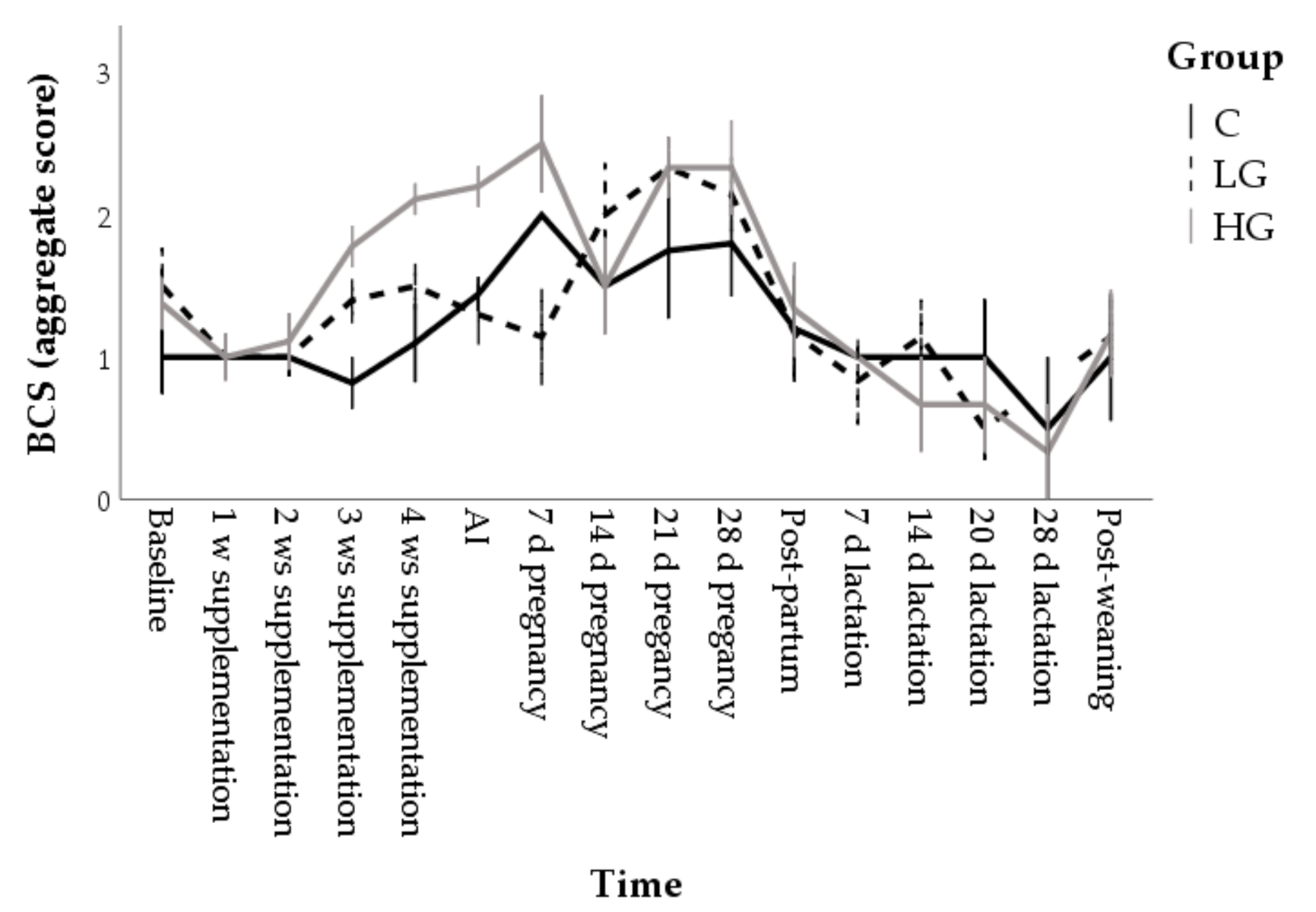
3.1.1. Body Conditions
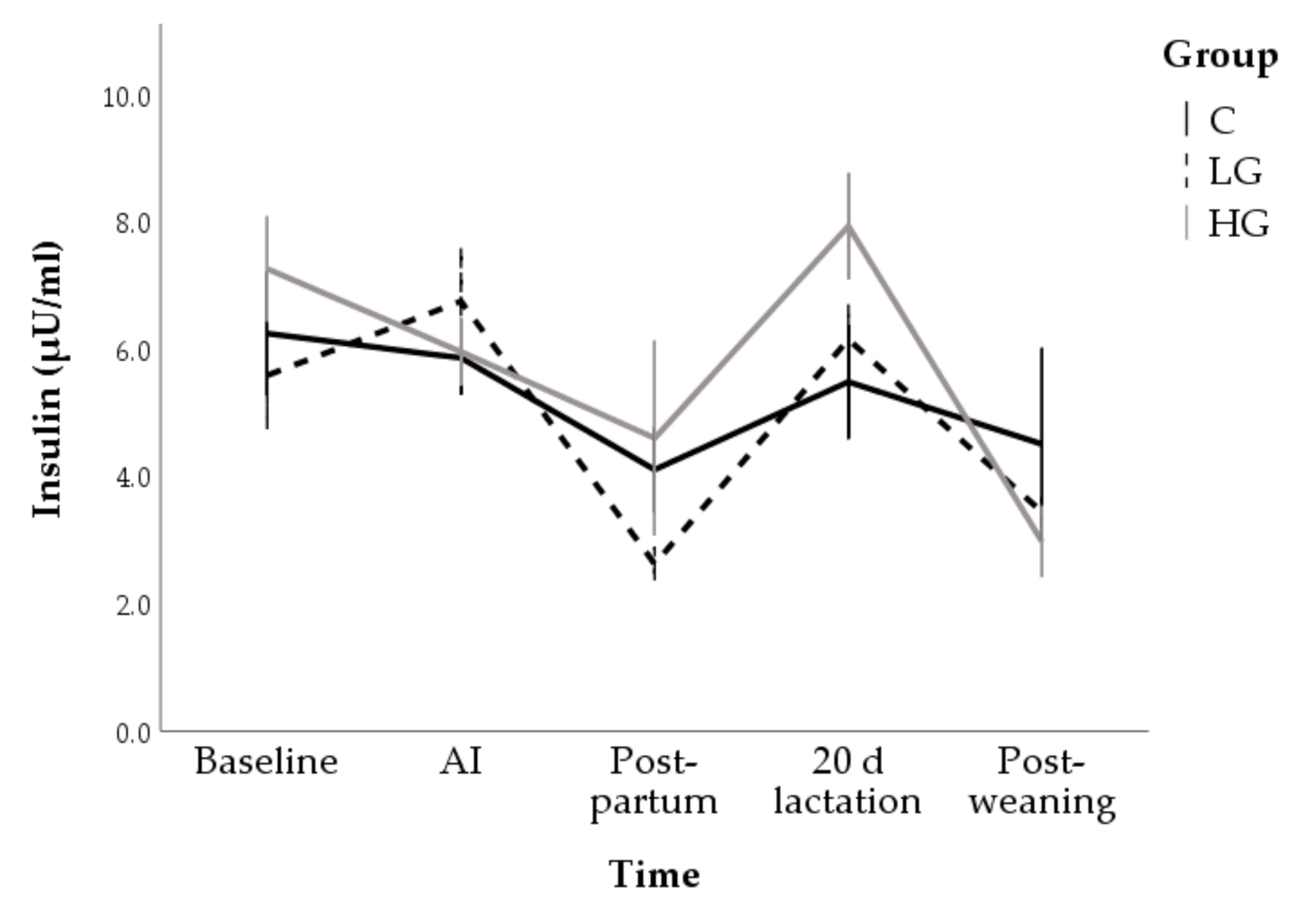
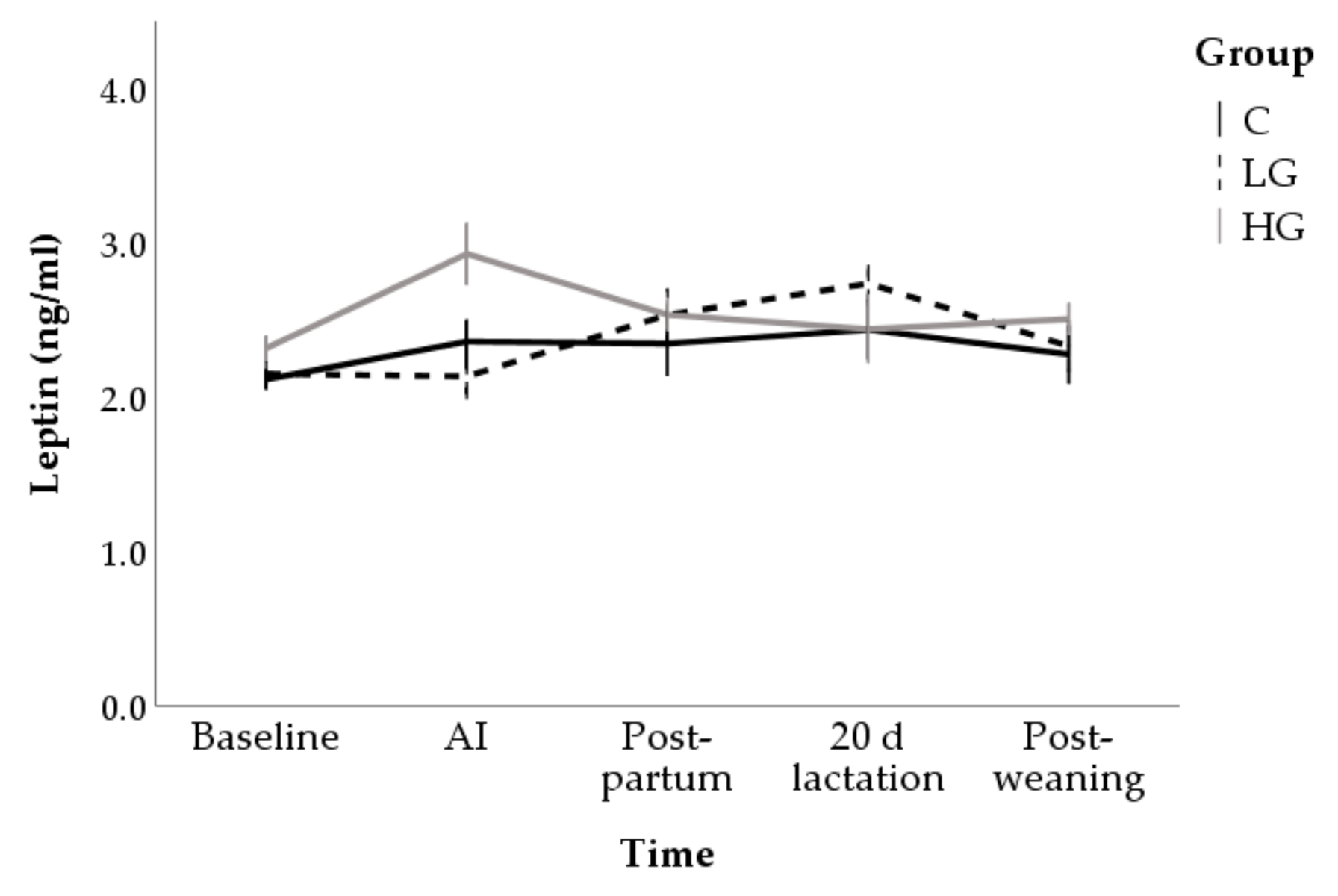
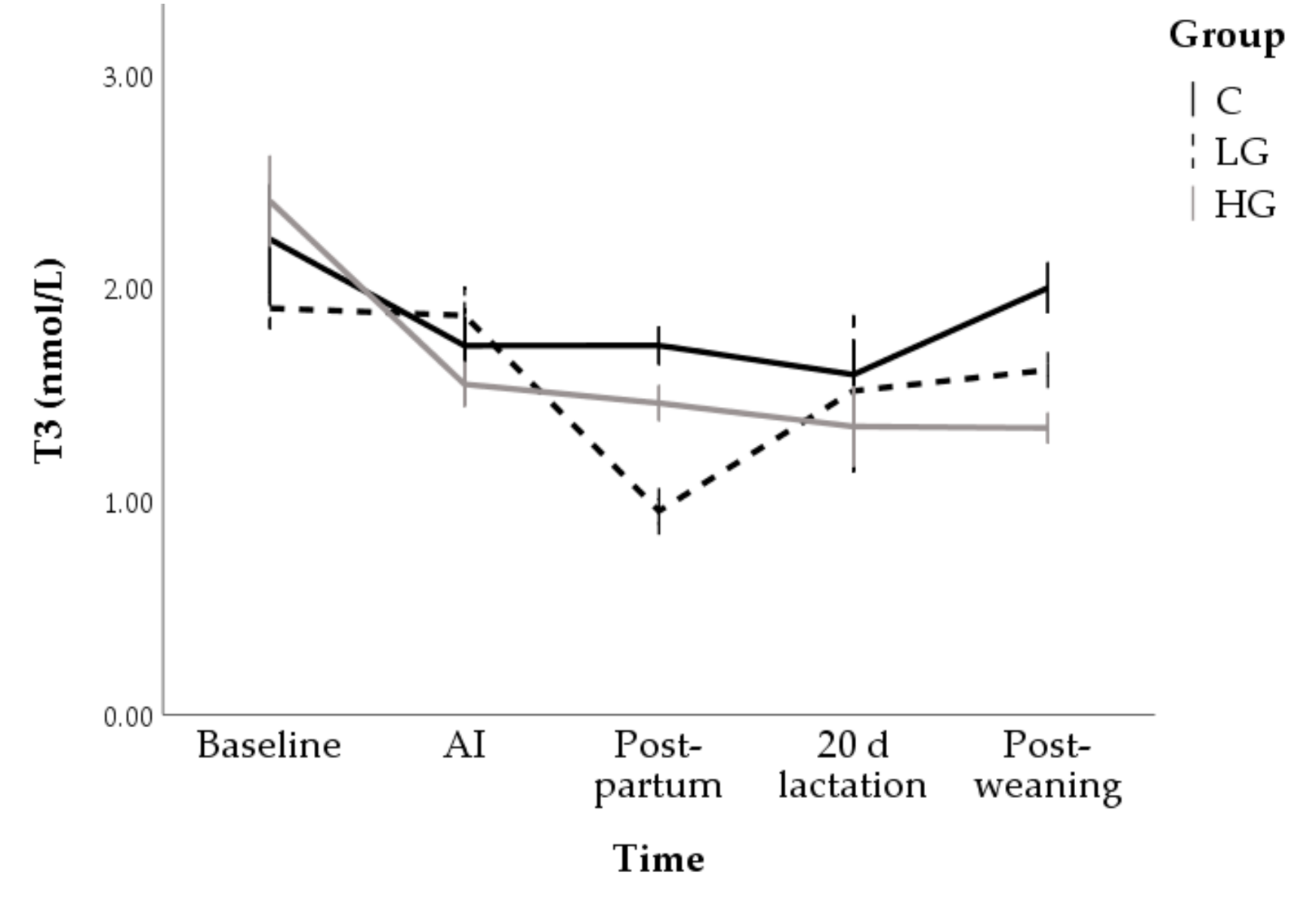
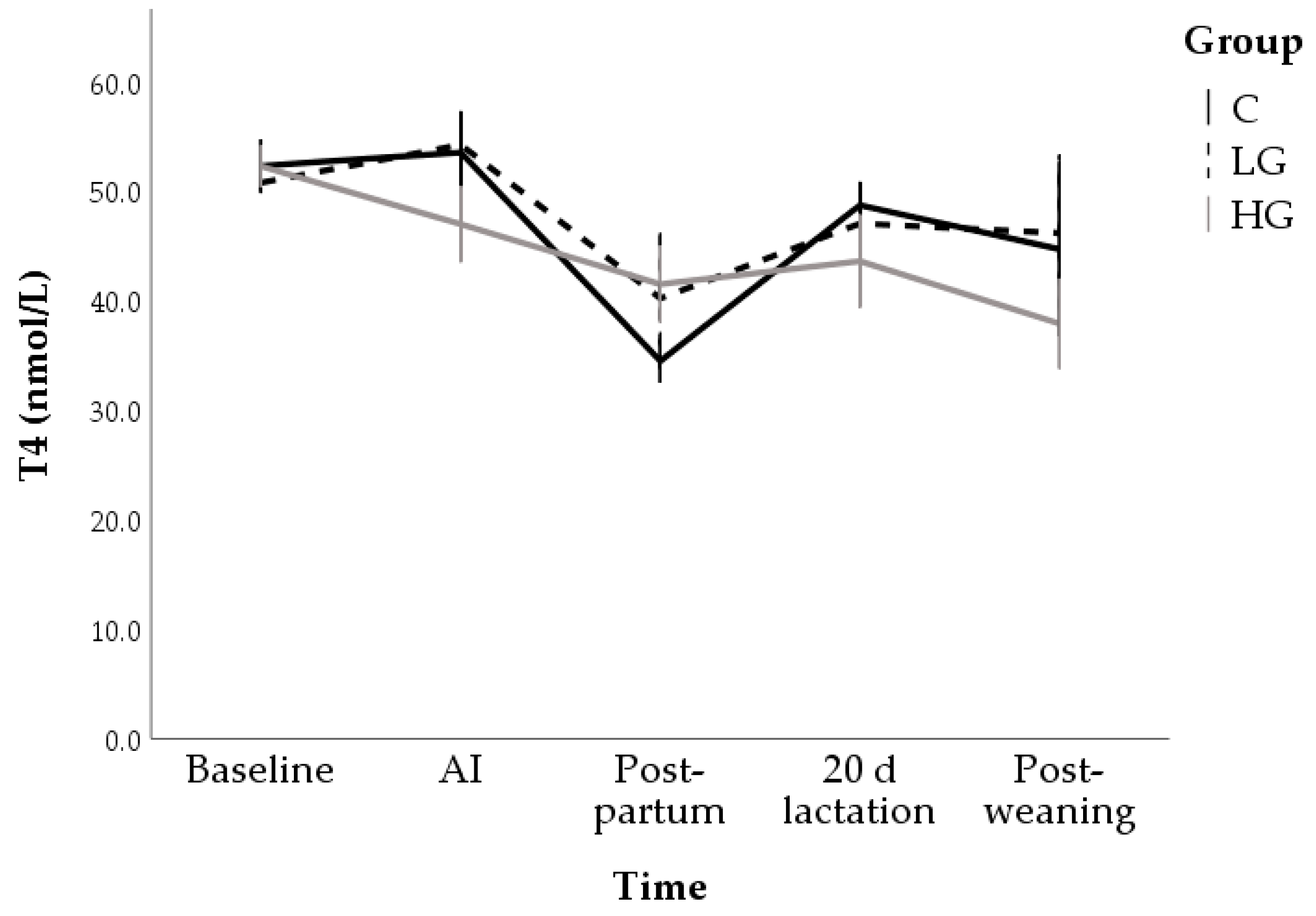
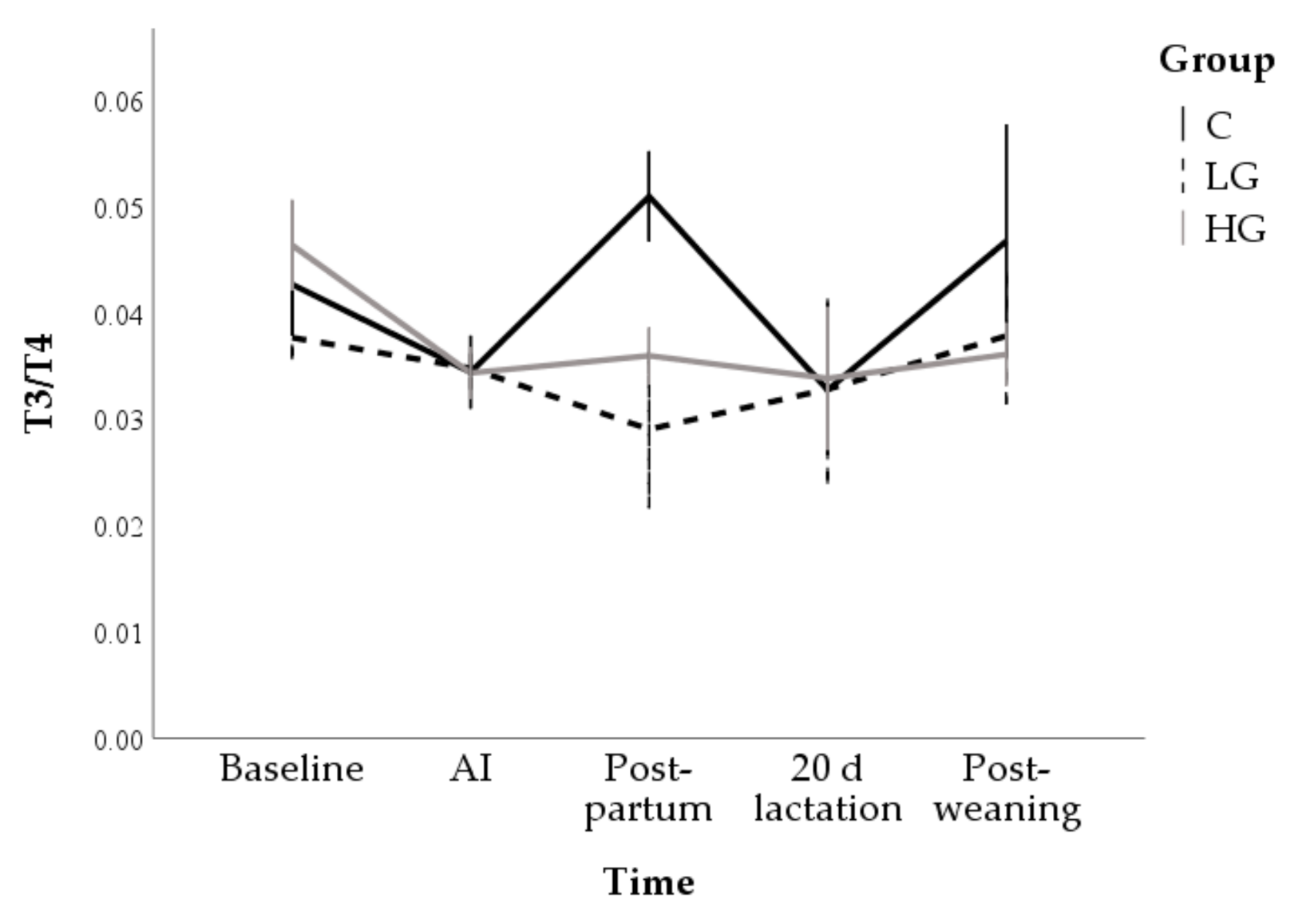
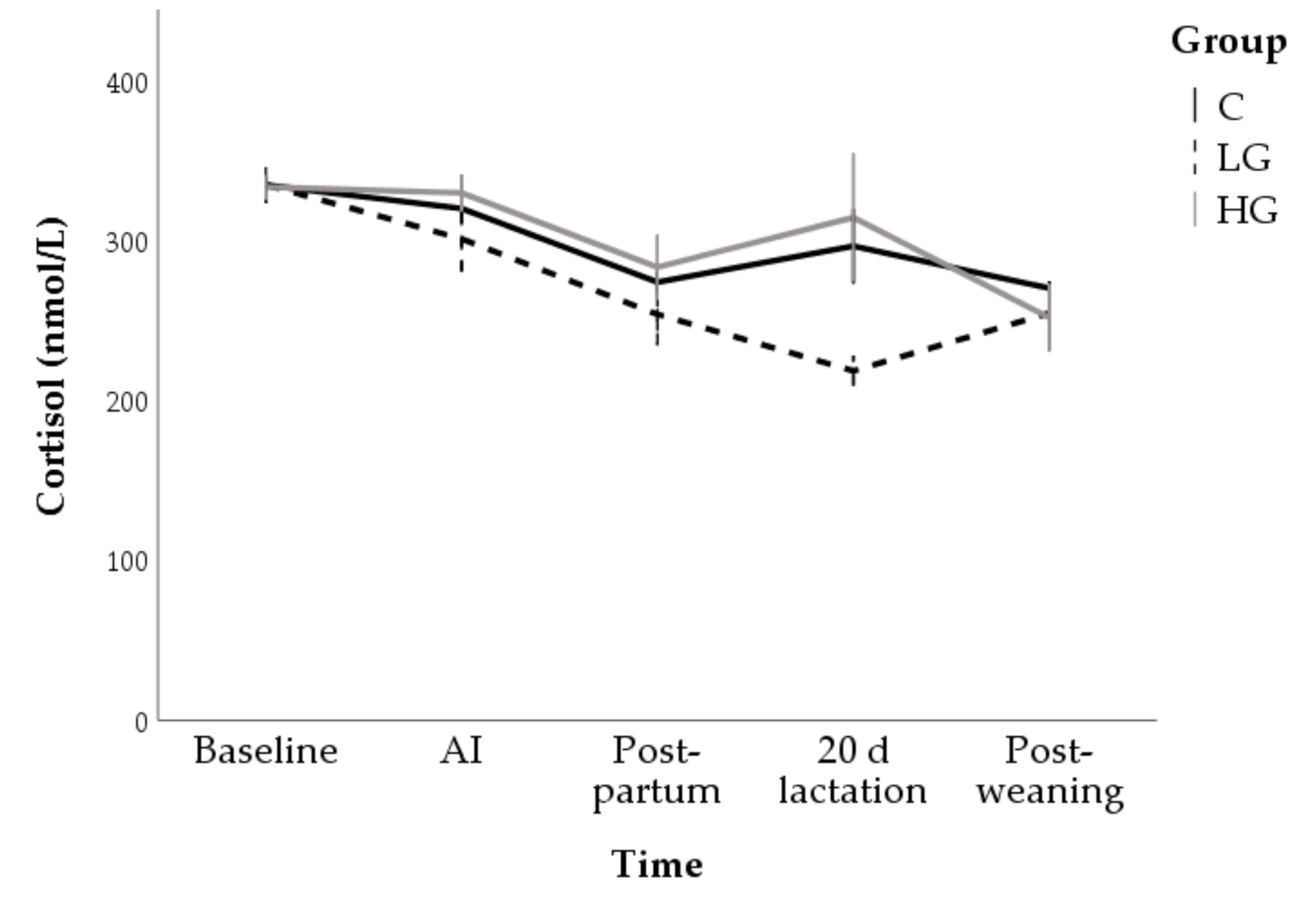
3.1.2. Hormone and Metabolite
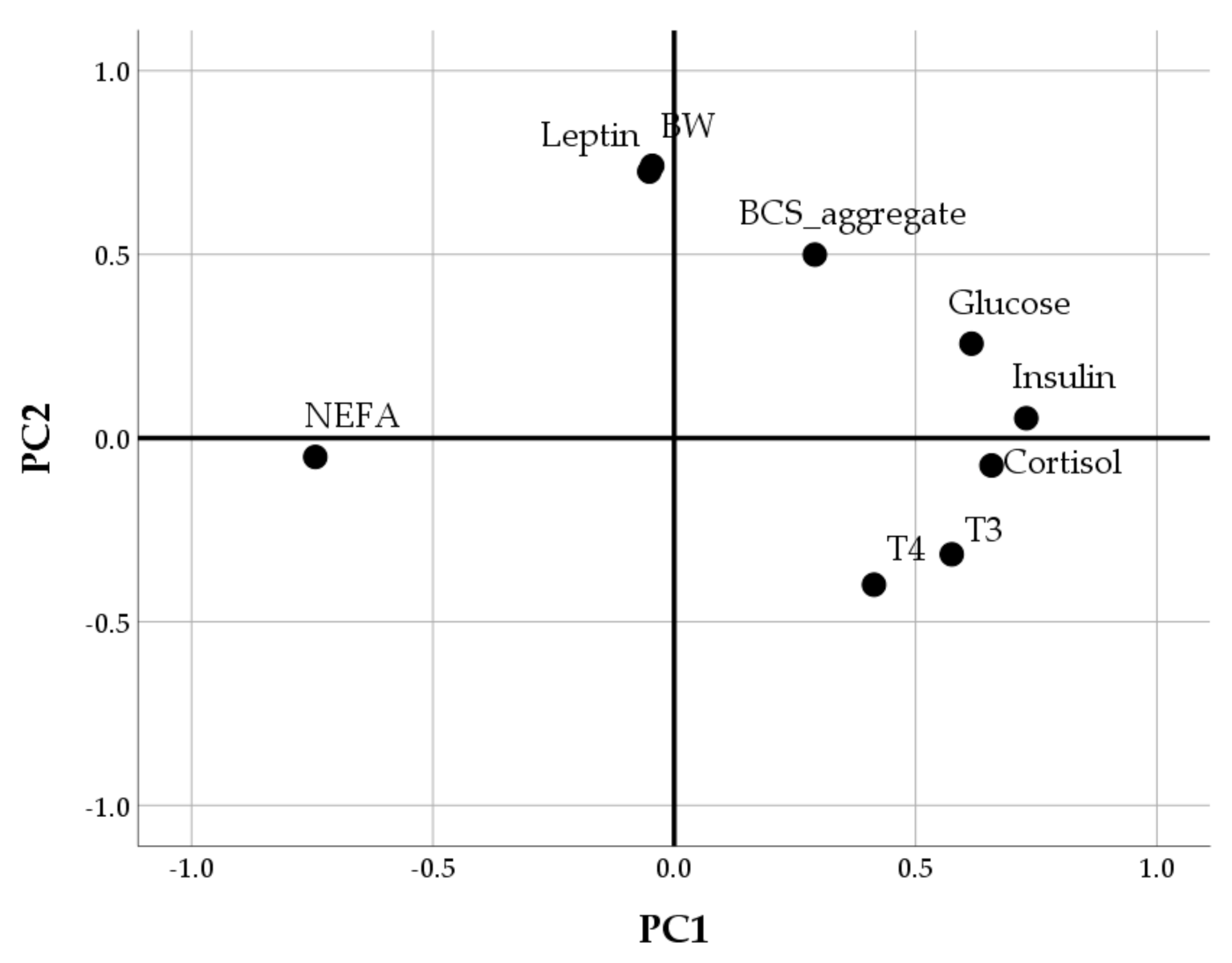
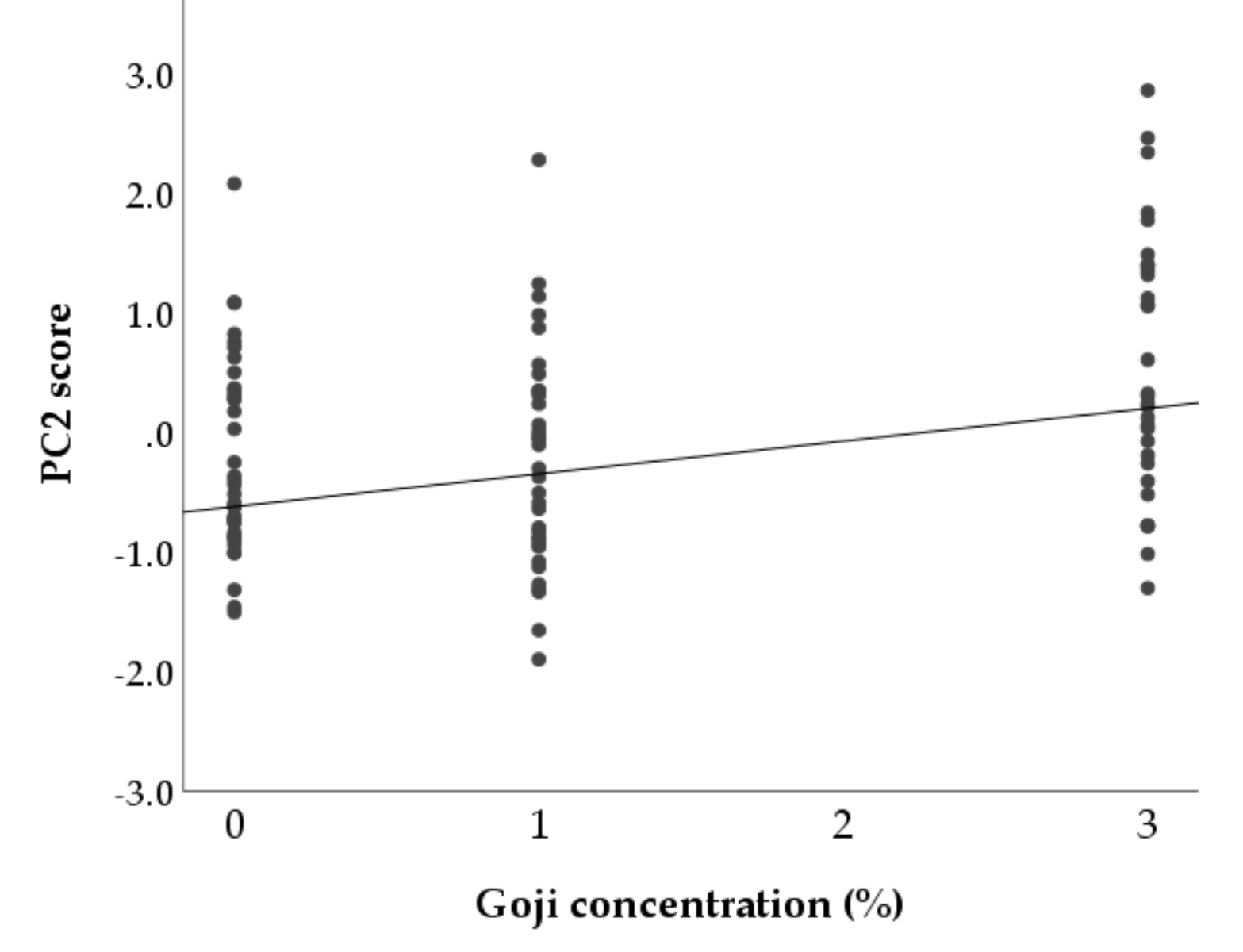
3.2. Multivariable Approach
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cheng, J.; Zhou, Z.-W.W.; Sheng, H.-P.P.; He, L.-J.J.; Fan, X.-W.W.; He, Z.-X.X.; Sun, T.; Zhang, X.; Zhao, R.J.; Gu, L.; et al. An evidence-based update on the pharmacological activities and possible molecular targets of Lycium barbarum polysaccharides. Drug Des. Devel. Ther. 2015, 9, 33–78. [Google Scholar] [PubMed] [Green Version]
- Potterat, O. Goji (Lycium barbarum and L. chinense): Phytochemistry, pharmacology and safety in the perspective of traditional uses and recent popularity. Planta Med. 2010, 76, 7–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.F.; Zhang, H.; Teh, S.S.; Wang, C.W.; Zhang, Y.; Hayford, F.; Wang, L.; Ma, T.; Dong, Z.; Zhang, Y.; et al. Goji Berries as a potential natural antioxidant medicine: An insight into their molecular mechanisms of action. Oxid. Med. Cell. Longev. 2019, 2019, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, H.; Liu, F.; Zuo, P.; Huang, G.; Song, Z.; Wang, T.; Lu, H.; Guo, F.; Han, C.; Sun, G. Practical application of antidiabetic efficacy of Lycium barbarum polysaccharide in patients with type 2 diabetes. Med. Chem. 2015, 11, 383–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgiev, K.D.; Slavov, I.J.; Iliev, I.A. Antioxidant activity and antiproliferative effects of Lycium barbarum’s (Goji berry) fractions on breast cancer cell lines. Folia Med. (Plovdiv) 2019, 61, 104–112. [Google Scholar] [CrossRef]
- Ceccarini, M.R.; Vannini, S.; Cataldi, S.; Moretti, M.; Villarini, M.; Fioretti, B.; Albi, E.; Beccari, T.; Codini, M. Effect of Lycium barbarum berries cultivated in Umbria (Italy) on human hepatocellular carcinoma cells. J. Biotechnol. 2016, 231, S26–S27. [Google Scholar] [CrossRef]
- You, J.; Chang, Y.; Zhao, D.; Zhuang, J.; Zhuang, W. A mixture of functional complex extracts from Lycium barbarum and grape seed enhances immunity synergistically in vitro and in vivo. J. Food Sci. 2019, 84, 1577–1585. [Google Scholar] [CrossRef]
- Zhao, R.; Li, Q.; Xiao, B. Effect of Lycium barbarum polysaccharide on the improvement of insulin resistance in NIDDM rats. Yakugaku Zasshi 2005, 125, 981–988. [Google Scholar] [CrossRef] [Green Version]
- Chien, K.J.; Horng, C.T.; Huang, Y.S.; Hsieh, Y.H.; Wang, C.J.; Yang, J.S.; Lu, C.C.; Chen, F.A. Effects of Lycium barbarum (goji berry) on dry eye disease in rats. Mol. Med. Rep. 2018, 17, 809–818. [Google Scholar] [CrossRef]
- Kang, Y.; Yang, G.; Zhang, S.; Ross, C.F.; Zhu, M.J. Goji berry modulates gut microbiota and alleviates colitis in IL-10-deficient mice. Mol. Nutr. Food Res. 2018, 62, e1800535. [Google Scholar] [CrossRef]
- Seeram, N. Berry fruits: Compositional elements, biochemical activities, and the impact of their intake on human health, performance, and disease. J. Agric. Food Chem. 2008, 56, 627–629. [Google Scholar] [CrossRef]
- Bucheli, P.; Vidal, K.; Shen, L.; Gu, Z.; Zhang, C.; Miller, L.E.; Wang, J. Goji berry effects on macular characteristics and plasma antioxidant levels. Optom. Vis. Sci. 2011, 88, 257–262. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Xiao, J.; Peng, B.; Xing, F.; So, K.-F.; Tipoe, G.L.; Lin, B. Retinal structure and function preservation by polysaccharides of wolfberry in a mouse model of retinal degeneration. Sci. Rep. 2014, 4, 7601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Q.; Li, J.; Yan, J.; Liu, S.; Guo, Y.; Chen, D.; Luo, Q. Lycium barbarum polysaccharides ameliorates renal injury and inflammatory reaction in alloxan-induced diabetic nephropathy rabbits. Life Sci. 2016, 157, 82–90. [Google Scholar] [CrossRef]
- Menchetti, L.; Vecchione, L.; Filipescu, I.; Petrescu, V.F.; Fioretti, B.; Beccari, T.; Ceccarini, M.R.; Codini, M.; Quattrone, A.; Trabalza-Marinucci, M.; et al. Effects of Goji berries supplementation on the productive performance of rabbit. Livest. Sci. 2019, 220, 123–128. [Google Scholar] [CrossRef]
- Menchetti, L.; Brecchia, G.; Branciari, R.; Barbato, O.; Fioretti, B.; Codini, M.; Bellezza, E.; Trabalza-Marinucci, M.; Miraglia, D. The effect of Goji berries (Lycium barbarum) dietary supplementation on rabbit meat quality. Meat Sci. 2020, 161, 108018. [Google Scholar] [CrossRef] [PubMed]
- Brecchia, G.; Menchetti, L.; Cardinali, R.; Castellini, C.; Polisca, A.; Zerani, M.; Maranesi, M.; Boiti, C. Effects of a bacterial lipopolysaccharide on the reproductive functions of rabbit does. Anim. Reprod. Sci. 2014, 147, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Paredes, E.; Ródenas, L.; Martínez-Vallespín, B.; Cervera, C.; Blas, E.; Brecchia, G.; Boiti, C.; Pascual, J.J. Effects of feeding programme on the performance and energy balance of nulliparous rabbit does. Animal 2012, 6, 1086–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boiti, C.; Guelfi, G.; Zerani, M.; Zampini, D.; Brecchia, G.; Gobbetti, A. Expression patterns of cytokines, p53 and nitric oxide synthase isoenzymes in corpora lutea of pseudopregnant rabbits during spontaneous luteolysis. Reproduction 2004, 127, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Foote, R.H.; Carney, E.W. The rabbit as a model for reproductive and developmental toxicity studies. Reprod. Toxicol. 2000, 14, 477–493. [Google Scholar] [CrossRef]
- Menchetti, L.; Barbato, O.; Sforna, M.; Vigo, D.; Mattioli, S.; Curone, G.; Tecilla, M.; Riva, F. Brecchia Effects of diets enriched in linseed and fish oil on the expression pattern of Toll-Like receptors 4 and proinflammatory cytokines on gonadal axis and reproductive organs in rabbit buck. Oxid. Med. Cell. Longev. 2020, 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menchetti, L.; Barbato, O.; Filipescu, I.E.; Traina, G.; Leonardi, L.; Polisca, A.; Troisi, A.; Guelfi, G.; Piro, F.; Brecchia, G. Effects of local lipopolysaccharide administration on the expression of Toll-like receptor 4 and pro-inflammatory cytokines in uterus and oviduct of rabbit does. Theriogenology 2018, 107, 162–174. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Xing, Y.; Qiao, C.; Liu, W.; Jiang, H.; Fu, Q.; Zhou, Y.; Yang, B.; Zhang, Z.; Chen, R. Semen quality improvement in boars fed with supplemental wolfberry (Lycium barbarum). Anim. Sci. J. 2019, 90, 1517–1522. [Google Scholar] [CrossRef]
- Bai, X.; Yan, X.; Xie, L.; Hu, X.; Lin, X.; Wu, C.; Zhou, N.; Wang, A.; See, M.T. Effects of pre-slaughter stressor and feeding preventative Chinese medicinal herbs on glycolysis and oxidative stability in pigs. Anim. Sci. J. 2016, 87, 1028–1033. [Google Scholar] [CrossRef] [PubMed]
- Castrica, M.; Menchetti, L.; Balzaretti, C.; Branciari, R.; Ranucci, D.; Cotozzolo, E.; Vigo, D.; Curone, G.; Brecchia, G.; Miraglia, D. Impact of dietary supplementation with goji berries (Lycium barbarum) on microbiological quality, physico-chemical, and sensory characteristics of rabbit meat. Foods 2020, 9, 1480. [Google Scholar] [CrossRef] [PubMed]
- Menchetti, L.; Brecchia, G.; Canali, C.; Cardinali, R.; Polisca, A.; Zerani, M.; Boiti, C. Food restriction during pregnancy in rabbits: Effects on hormones and metabolites involved in energy homeostasis and metabolic programming. Res. Vet. Sci. 2015, 98, 7–12. [Google Scholar] [CrossRef]
- Menchetti, L.; Canali, C.; Castellini, C.; Boiti, C.; Brecchia, G. The different effects of linseed and fish oil supplemented diets on insulin sensitivity of rabbit does during pregnancy. Res. Vet. Sci. 2018, 118, 126–133. [Google Scholar] [CrossRef]
- Menchetti, L.; Brecchia, G.; Cardinali, R.; Polisca, A.; Boiti, C. Feed restriction during pregnancy: Effects on body condition and productive performance of primiparous rabbit does. World Rabbit Sci. 2015, 23, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Fortun-Lamothe, L. Energy balance and reproductive performance in rabbit does. Anim. Reprod. Sci. 2006, 93, 1–15. [Google Scholar] [CrossRef]
- Troisi, A.; Cardinali, L.; Menchetti, L.; Speranza, R.; Verstegen, J.P.; Polisca, A. Serum concentrations of leptin in pregnant and non-pregnant bitches. Reprod. Domest. Anim. 2020, 55, 454–459. [Google Scholar] [CrossRef]
- Menchetti, L.; Andoni, E.; Barbato, O.; Canali, C.; Quattrone, A.; Vigo, D.; Codini, M.; Curone, G.; Brecchia, G. Energy homeostasis in rabbit does during pregnancy and pseudopregnancy. Anim. Reprod. Sci. 2020, 218, 106505. [Google Scholar] [CrossRef] [PubMed]
- Ladyman, S.R.; Augustine, R.A.; Grattan, D.R. Hormone interactions regulating energy balance during pregnancy. J. Neuroendocrinol. 2010, 22, 805–817. [Google Scholar] [CrossRef] [PubMed]
- Brecchia, G.; Bonanno, A.; Galeati, G.; Federici, C.; Maranesi, M.; Gobbetti, A.; Zerani, M.; Boiti, C. Hormonal and metabolic adaptation to fasting: Effects on the hypothalamic-pituitary-ovarian axis and reproductive performance of rabbit does. Domest. Anim. Endocrinol. 2006, 31, 105–122. [Google Scholar] [CrossRef] [PubMed]
- Maertens, L.; Moermans, R.; De Groote, G. Prediction of the apparent digestible energy content of commercial pelleted feeds for rabbits. J. Appl. Rabbit Res. 1988, 11, 60–67. [Google Scholar]
- García-García, R.M.; Rebollar, P.G.; Arias-Álvarez, M.; Sakr, O.G.; Bermejo-Álvarez, P.; Brecchia, G.; Gutierrez-Adan, A.; Zerani, M.; Boit, C.; Lorenzo, P.L. Acute fasting before conception affects metabolic and endocrine status without impacting follicle and oocyte development and embryo gene expression in the rabbit. Reprod. Fertil. Dev. 2011, 23, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Garson, G.D. Factor Analysis; Statistical Associates Publishers: Asheboro, NC, USA, 2013. [Google Scholar]
- Righi, C.; Menchetti, L.; Orlandi, R.; Moscati, L.; Mancini, S.; Diverio, S. Welfare Assessment in Shelter Dogs by Using Physiological and Immunological Parameters. Animals 2019, 9, 340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agradi, S.; Curone, G.; Negroni, D.; Vigo, D.; Brecchia, G.; Bronzo, V.; Chiesa, L.; Peric, T.; Danes, D.; Menchetti, L. Determination of fatty acids profile in Original Brown cows dairy products and relationship with alpine pasture farming system. Animals 2020, 10, 1231. [Google Scholar] [CrossRef]
- Menchetti, L.; Calipari, S.; Guelfi, G.; Catanzaro, A.; Diverio, S. My dog is not my cat: Owner perception of the personalities of dogs and cats living in the same household. Animals 2018, 8, 80. [Google Scholar] [CrossRef] [Green Version]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioural Sciences, 2nd ed.; Lawrence Erlbaum Associates Inc.: Mahwah, NJ, USA, 1988. [Google Scholar]
- Kawai, M.; Yamaguchi, M.; Murakami, T.; Shima, K.; Murata, Y.; Kishi, K. The placenta is not the main source of leptin production in pregnant rat: Gestational profile of leptin in plasma and adipose tissues. Biochem. Biophys. Res. Commun. 1997, 240, 798–802. [Google Scholar] [CrossRef]
- Block, S.S.; Butler, W.R.; Ehrhardt, R.A.; Bell, A.W.; Van Amburgh, M.E.; Boisclair, Y.R. Decreased concentration of plasma leptin in periparturient dairy cows is caused by negative energy balance. J. Endocrinol. 2001, 171, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, L.; Troisi, A.; Verstegen, J.P.; Menchetti, L.; Elad Ngonput, A.; Boiti, C.; Canello, S.; Zelli, R.; Polisca, A. Serum concentration dynamic of energy homeostasis hormones, leptin, insulin, thyroid hormones, and cortisol throughout canine pregnancy and lactation. Theriogenology 2017, 97, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Ciampelli, M.; Lanzone, A.; Caruso, A. Insulin in obstetrics: A main parameter in the management of pregnancy. Hum. Reprod. Update 1998, 4, 904–914. [Google Scholar] [CrossRef] [Green Version]
- Maertens, L.; Lebas, F.; Szendrö, Z. Rabbit milk: A review of quantity, quality and non-dietary affecting factors. World Rabbit Sci. 2006, 14, 205–230. [Google Scholar] [CrossRef]
- Baruselli, P.S.; Vieira, L.M.; Sá Filho, M.F.; Mingoti, R.D.; Ferreira, R.M.; Chiaratti, M.R.; Oliveira, L.H.; Sales, J.N.; Sartori, R. Associations of insulin resistance later in lactation on fertility of dairy cows. Theriogenology 2016, 86, 263–269. [Google Scholar] [CrossRef]
- Leiva, T.; Cooke, R.F.; Brandão, A.P.; Aboin, A.C.; Ranches, J.; Vasconcelos, J.L.M. Effects of excessive energy intake and supplementation with chromium propionate on insulin resistance parameters, milk production, and reproductive outcomes of lactating dairy cows. Livest. Sci. 2015, 180, 121–128. [Google Scholar] [CrossRef] [Green Version]
- De Koster, J.D.; Opsomer, G. Insulin resistance in dairy cows. Vet. Clin. N. Am. Food Anim. Pract. 2013, 29, 299–322. [Google Scholar] [CrossRef] [PubMed]
- Leblanc, S. Monitoring metabolic health of dairy cattle in the transition period. J. Reprod. Dev. 2010, 56, S29–S35. [Google Scholar] [CrossRef] [Green Version]
- Bossaert, P.; Leroy, J.L.M.R.; De Vliegher, S.; Opsomer, G. Interrelations between glucose-induced insulin response, metabolic indicators, and time of first ovulation in high-yielding dairy cows. J. Dairy Sci. 2008, 91, 3363–3371. [Google Scholar] [CrossRef]
- Munir, K.M.; Chandrasekaran, S.; Gao, F.; Quon, M.J. Mechanisms for food polyphenols to ameliorate insulin resistance and endothelial dysfunction: Therapeutic implications for diabetes and its cardiovascular complications. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E679–E686. [Google Scholar] [CrossRef] [Green Version]
- Caimari, A.; Mariné-Casadó, R.; Boqué, N.; Crescenti, A.; Arola, L.; Del Bas, J.M. Maternal intake of grape seed procyanidins during lactation induces insulin resistance and an adiponectin resistance-like phenotype in rat offspring. Sci. Rep. 2017, 7, 12573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Jiang, G.; Kebreab, E.; Yu, Q.; Li, J.; Zhang, X.; He, H.; Fang, R.; Dai, Q. Effects of dietary grape seed polyphenols supplementation during late gestation and lactation on antioxidant status in serum and immunoglobulin content in colostrum of multiparous sows. J. Anim. Sci. 2019, 97, 2515–2523. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhao, L.; Gao, M.; Loor, J.J. Tea polyphenols protect bovine mammary epithelial cells from hydrogen peroxide-induced oxidative damage in vitro. J. Anim. Sci. 2018, 96, 4159–4172. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, S.; Norikura, T.; Sato, S. Maternal green tea polyphenol intake during lactation attenuates kidney injury in high-fat-diet-fed male offspring programmed by maternal protein restriction in rats. J. Nutr. Biochem. 2018, 56, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Kita, T.; Yamasaki, S.; Kawahara, T.; Ueno, Y.; Yamada, M.; Mukai, Y.; Sato, S.; Kurasaki, M.; Saito, T. Maternal resveratrol intake during lactation attenuates hepatic triglyceride and fatty acid synthesis in adult male rat offspring. Biochem. Biophys. Reports 2017, 9, 173–179. [Google Scholar] [CrossRef]
- Castellini, C.; Dal Bosco, A.; Cardinali, R. Long term effect of post-weaning rhythm on the body fat and performance of rabbit doe. Reprod. Nutr. Dev. 2006, 46, 195–204. [Google Scholar] [CrossRef]
- Da Costa, T.H.M.; Bluck, L.J. High lactation index is associated with insulin sensitivity. J. Nutr. Biochem. 2011, 22, 446–449. [Google Scholar] [CrossRef]
- Ahima, R.S.; Osei, S.Y. Leptin signaling. Physiol. Behav. 2004, 81, 223–241. [Google Scholar] [CrossRef]
- Johnstone, L.E.; Higuchi, T. Food intake and leptin during pregnancy and lactation. Prog. Brain Res. 2001, 133, 215–227. [Google Scholar]
- Vernon, R.G.; Denis, R.G.P.; Sorensen, A.; Williams, G. Leptin and the adaptations of lactation in rodents and ruminants. Horm. Metab. Res. 2002, 34, 678–685. [Google Scholar] [CrossRef]
- Casabiell, X.; Piñeiro, V.; Tomé, M.A.; Peinó, R.; Diéguez, C.; Casanueva, F.F. Presence of leptin in colostrum and/or breast milk from lactating mothers: A potential role in the regulation of neonatal food intake. J. Clin. Endocrinol. Metab. 1997, 82, 4270–4273. [Google Scholar] [CrossRef] [PubMed]
- Palou, M.; Picó, C.; Palou, A. Leptin as a breast milk component for the prevention of obesity. Nutr. Rev. 2018, 76, 875–892. [Google Scholar] [CrossRef] [PubMed]
- Picó, C.; Oliver, P.; Sánchez, J.; Miralles, O.; Caimari, A.; Priego, T.; Palou, A. The intake of physiological doses of leptin during lactation in rats prevents obesity in later life. Int. J. Obes. 2007, 31, 1199–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, E.; Hue-Beauvais, C.; Galio, L.; Solomon, G.; Gertler, A.; Revillon, F.; Lhotellier, V.; Aujean, E.; Devinoy, E.; Charlier, M. Leptin gene in rabbit: Cloning and expression in mammary epithelial cells during pregnancy and lactation. Physiol. Genomics 2013, 45, 645–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrannini, E.; Camastra, S.; Coppack, S.; Fliser, D.; Golay, A.; Mitrakou, A. Insulin action and non-esterified fatty acids. Proc. Nutr. Soc. 1997, 56, 753–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Unit | Diet | ||
|---|---|---|---|---|
| C | LG | HG | ||
| Ingredients | ||||
| Wheat bran | % | 30.0 | 29.5 | 29.0 |
| Dehydrated alfalfa meal | % | 42.0 | 41.5 | 41.0 |
| Barley | % | 9.5 | 9.5 | 9.0 |
| Sunflower meal | % | 4.5 | 4.5 | 4.2 |
| Rice bran | % | 4.0 | 4.0 | 3.9 |
| Soybean meal | % | 4.0 | 4.0 | 3.9 |
| Calcium carbonate | % | 2.2 | 2.2 | 2.2 |
| Cane molasses | % | 2.0 | 2.0 | 2.0 |
| Vitamin-mineral premix * | % | 0.4 | 0.4 | 0.4 |
| Soybean oil | % | 0.4 | 0.4 | 0.4 |
| Salt | % | 0.3 | 0.3 | 0.3 |
| Goji berries | % | - | 1.0 | 3.0 |
| Analytical data | ||||
| Crude Protein | % | 15.74 | 15.64 | 15.66 |
| Ether extract | % | 2.25 | 2.23 | 2.47 |
| Ash | % | 9.28 | 9.36 | 9.25 |
| Starch | % | 16.86 | 17.07 | 16.99 |
| NDF | % | 38.05 | 38.55 | 37.49 |
| ADF | % | 19.54 | 19.60 | 19.01 |
| ADL | % | 4.01 | 4.31 | 3.98 |
| Digestible Energy ** | Kcal/kg | 2464 | 2463 | 2459 |
| Item | Component | |
|---|---|---|
| PC1 | PC2 | |
| NEFA | −0.744 | −0.051 |
| Insulin | 0.729 | 0.054 |
| Cortisol | 0.658 | −0.075 |
| Glucose | 0.616 | 0.257 |
| T3 | 0.575 | −0.316 |
| T4 | 0.414 | −0.399 |
| BW | −0.046 | 0.741 |
| Leptin | −0.052 | 0.725 |
| BCS | 0.291 | 0.499 |
| % Variance explained | 27.7% | 18.4% |
| Cumulative % variance explained | 46.1% | |
| Dependent Variable/Outcome | Independent Variable/Predictor | B Coefficient | Standard Error | p Value |
|---|---|---|---|---|
| PC1 | Constant | 0.611 | 0.1511 | <0.001 |
| Goji berries concentration in feed | 0.019 | 0.0663 | 0.776 | |
| Days of supplementation | −0.017 | 0.0024 | <0.001 | |
| PC2 | Constant | −0.619 | 0.1373 | <0.001 |
| Goji berries concentration in feed | 0.275 | 0.0765 | <0.001 | |
| Days of supplementation | 0.007 | 0.0027 | 0.006 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menchetti, L.; Curone, G.; Andoni, E.; Barbato, O.; Troisi, A.; Fioretti, B.; Polisca, A.; Codini, M.; Canali, C.; Vigo, D.; et al. Impact of Goji Berries (Lycium barbarum) Supplementation on the Energy Homeostasis of Rabbit Does: Uni- and Multivariate Approach. Animals 2020, 10, 2000. https://doi.org/10.3390/ani10112000
Menchetti L, Curone G, Andoni E, Barbato O, Troisi A, Fioretti B, Polisca A, Codini M, Canali C, Vigo D, et al. Impact of Goji Berries (Lycium barbarum) Supplementation on the Energy Homeostasis of Rabbit Does: Uni- and Multivariate Approach. Animals. 2020; 10(11):2000. https://doi.org/10.3390/ani10112000
Chicago/Turabian StyleMenchetti, Laura, Giulio Curone, Egon Andoni, Olimpia Barbato, Alessandro Troisi, Bernard Fioretti, Angela Polisca, Michela Codini, Claudio Canali, Daniele Vigo, and et al. 2020. "Impact of Goji Berries (Lycium barbarum) Supplementation on the Energy Homeostasis of Rabbit Does: Uni- and Multivariate Approach" Animals 10, no. 11: 2000. https://doi.org/10.3390/ani10112000