|
|
In: Van Driesche, R., et al., 2002, Biological Control of Invasive Plants in the Eastern United States, USDA Forest Service Publication FHTET-2002-04, 413 p.
moister sites. Canada thistle was among the most prevalent weeds on Conservation Reserve Program (CRP) land in Minnesota, occurring in 65 to 75% of CRP fields throughout the state. Canada thistle ground cover in these fields frequently reached 50 to 75%, giving rise to concern about seed dispersal into neighboring agricultural land (Jewett et al., 1996). It was ranked as “urgent“ for control in a review of exotic plants at Pipestone National Monument, Minnesota (Hiebert and Stubbendieck, 1993).
Extent of losses. A density of 20 Canada thistle shoots per m2 caused estimated yield losses of 34% in barley (O’Sullivan et al., 1982), 26% in canola (O’Sullivan et al., 1985), 36% in winter wheat (McLennan et al., 1991), and 48% in alfalfa seed (Moyer et al., 1991). Densities of Canada thistle in field infestations can reach 173 shoots per m2 (Donald and Khan, 1996). Geographical Distribution Canada thistle occurs in all eastern U.S. states south to Kansas, Arkansas, Tennessee, and North Carolina, but it is sparsely distributed south of latitude 37°N (USDA, NRCS, 1999). The main areas of occurrence are the northeastern, mid-Atlantic, Great Lakes, and northern Great Plains states. In a survey in Maryland, Canada thistle was found in about 17% of suitable sites in the eastern and central part of the state, but only 10% of sites further west (Tipping, 1992). Background Information On The Pest Plant Taxonomy Canada thistle is a member of the genus Cirsium, subtribe Carduinae, tribe Cardueae, and family Asteraceae (Bremer, 1994). It differs from most other Cirsium species by its dioecious flowers, and from most native North American members of the genus by its extensive creeping roots and small, numerous flower heads borne on branched stems. Several varieties have been described based on variations in leaf shape and degree of spininess. Biology The biology of Canada thistle was extensively reviewed by Moore (1975), Donald (1994), and Nuzzo (1997). It is a perennial herb with an extensive creeping root system that can give rise to new shoots from adventitious root buds. The stems usually die back over winter and new shoots are produced each spring from old stem bases or root buds. Canada thistle is almost perfectly dioecious and can produce abundant seeds, which are dispersed by wind (Lloyd and Myall, 1976). It is a long-day plant, requiring a photoperiod of at least 14 to 16 hours (depending on ecotype) for flowering to be induced (Hunter and Smith, 1972). It occurs in a wide range of habitats and soil types. Analysis of Related Native Plants in the Eastern United States The genus Cirsium is a large one, with 92 native species in North America, of which 20 occur in the U.S. states that fall wholly or in part east of the 100th meridian (USDA, NRCS, 1999). One of these, Cirsium pitcheri (Torrey) Torrey and Gray, is listed as threatened under the Endangered Species Act. This species occurs in sand dunes along the shores of the Great Lakes in Illinois, Indiana, Michigan, Wisconsin, and Ontario. Phylogenetic studies of North American and Eurasian Cirsium species are needed to elucidate relationships among species in the genus and to provide a basis for planning host-specificity tests and interpreting resulting data. Studies have been initiated using the external transcribed spacer (ETS) region of ribosomal DNA to develop a phylogeny for North American and selected Eurasian Cirsium species (D. Kelch, pers. comm.). History of Biological Control Efforts in the Eastern United States Canada thistle was among the first 19 weed species selected as targets for biological control when the USDA Rome Laboratory was established in 1959 (Schroeder, 1980). However, most host specificity testing of agents for Canada thistle was conducted from 1961 to 1984 by staff of Agriculture Canada or by the International Institute of Biological Control (now CABI Bioscience) working with Canadian funding. The agents released in the United States have been those that became available as a result of the Canadian program, the results of which were reviewed by Schroeder (1980), Peschken (1984a), and McClay et al. (2001). Most releases in the eastern United States were made by USDA, ARS staff at the Beltsville Agricultural Research Center; some studies also were carried out by staff of the Maryland Department of Agriculture. Biological control of Canada thistle in New Zealand has been reviewed by Jessep (1989). Area of Origin of Weed Canada thistle is native to Europe, parts of North Africa, and Asia south to Afghanistan, Iran and Pakistan, and east to China. Its exact center of origin within the native range is not known, although it is suggested by Moore (1975) to be in southeastern Europe and the eastern Mediterranean area. Areas Surveyed for Natural Enemies Extensive surveys of natural enemies of Canada thistle and other Cardueae species in western Europe were carried out starting in 1959. Other surveys have been carried out in Japan, Iran, and northern Pakistan (Schroeder, 1980), and in China (Wan and Harris, unpub. data). Further surveys in southern Russia, central Asia, and China are currently under way (Gassmann, unpub. data). In addition to surveys specifically carried out for biocontrol purposes, the general European entomological literature contains much information on insects associated with Canada thistle (e.g., Redfern, 1983; Stary, 1986; Volkl, 1989; Freese, 1994; Berestetsky, 1997; Frenzel et al., 2000). The phytophagous insects associated with Canada thistle in Poland are listed by Winiarska (1986). Natural Enemies Found Surveys by Zwölfer (1965a) in Europe found 78 species of phytophagous insects feeding on Canada thistle. Of these, six are reportedly monophagous, five are found on Canada thistle and a few related species, 26 are oligophagous on plants in the same subtribe, and the remaining 42 are less specific and of no interest for biological control (Schroeder, 1980). A number of European insects and pathogens attacking Canada thistle have been accidentally introduced into North America, and some of these have been studied as potential biological control agents. The leaf-feeding tortoise beetle Cassida rubiginosa Müller (Coleoptera: Chrysomelidae) occurs widely in the eastern United States, south to Virginia and west to southern Michigan and Ohio, and in Canada (Ward and Pienkowski, 1978a). The seed-feeding weevil Larinus planus (F.) (Coleoptera: Curculionidae) was found by Wheeler and Whitehead (1985) to be well established in Pennsylvania, Maryland, Ohio, and New York, with the earliest records dating from 1968 in Ohio. It has also been collected from Indiana and West Virginia (C. W. O‘Brien, pers. comm.). The seed-head fly Terellia ruficauda (F.) (=Orellia ruficauda F.) (Diptera: Tephritidae) is distributed across Canada, and presumably also occurs widely in the eastern United States. A survey showed it to be present in South Dakota (R. Moehringer, S. Dakota Dept. of Agriculture, pers. comm.), and specimens are known from Michigan. The root-feeding weevil Cleonis pigra (Scopoli) (Coleoptera: Curculionidae) occurs in New York, Pennsylvania, Michigan, Indiana, Ontario, and Quebec (O’Brien and Wibmer, 1982; Anderson, 1987; C. W. O‘Brien, per. comm.). The rust Puccinia punctiformis (Strauss) Röhling is widespread in North America. A phytopathogenic bacterium, Pseudomonas syringae pv. tagetis (Hellmers 1955) Young, Dye and Wilkie 1978, causing apical chlorosis, has been isolated from Canada thistle. Field tests of applications of this bacterium in a commercial corn field resulted in 57% mortality of Canada thistle as well as damage to several other weedy Asteraceae species. A surfactant is required to allow penetration of the Canada thistle cuticle by the bacterium. Further work on formulation of this agent is under way (Johnson et al., 1996). The bacterium occurs in Maryland (P. Tipping, pers. comm.). One species which was introduced as a biological control agent for Carduus species, the seed-head weevil Rhinocyllus conicus (Frölich) (Coleoptera: Curculionidae), also is recorded attacking Canada thistle (Rees, 1977; Youssef and Evans, 1994). This species is widespread in the eastern United States, and has been found attacking Canada thistle in Maryland (P. Tipping, pers. comm.). Host Range Tests and Results In the earlier part of the period 1961 to 1984, host specificity testing for agents attacking Canada thistle was focused on assessing potential risks to economic species of Cardueae, of which the two most important are safflower (Carthamus tinctorius L.) and globe artichoke (Cynara scolymus L.). In later studies, some native North American Cirsium species also were tested, but potential impacts of most agents on native non-target Cirsium species were not assessed in detail. The leaf-feeding beetle Altica carduorum Guérin-Méneville (Coleoptera: Chrysomelidae) is known in the field in Europe mainly from Canada thistle, with a single record of adults from Carduus pycnocephalus L. (Zwölfer, 1965a). The host specificity of a population of A. carduorum from Switzerland was studied by Harris (1964), using starvation tests with adults and larvae. First instar larvae complete development only on Cirsium, Carduus, and Silybum species. Adults feed readily on all Cirsium species tested, which included only two North American species, but their feeding rate is highest on Canada thistle. Similar results were obtained by Karny (1963) and Zwölfer (1965b). More recently, the host specificity of a biotype of A. carduorum from Xinjiang, China, was assessed by Wan et al. (1996), who found that in no-choice tests this beetle can complete development on 18 Cirsium species (mostly North American) and Silybum marianum (L.) Gaertner. A risk analysis approach, however, predicted that North American Cirsium species would be safe from attack in the field because host selection requires a series of sequential steps, with the native species being less preferred than C. arvense at each stage (Wan and Harris, 1997). It also was suggested that the insect is monophagous in the field because host finding is dependent on aggregation to substances from wounds and feces specific to C. arvense (Wan and Harris, 1996). As the Xinjiang biotype was not approved for field release in North America, it has not been possible to test these predictions in the field. The only known field host plant of the weevil Ceutorhynchus litura (F.) (Coleoptera: Curculionidae) in Europe is Canada thistle, except for three collections from Carduus defloratus L. in Switzerland (Zwölfer and Harris, 1966). Ceutorhynchus litura was screened by Zwölfer and Harris (1966), who found that feeding, oviposition, and larval development are restricted to species in the genera Cirsium, Carduus, and Silybum. Normal larval development occurs on all Cirsium species tested, including three native North American species. Ceutorhynchus litura was approved for release in Canada and the United States based on its lack of attack on economic Cardueae species. In a more recent European field survey of seven Cirsium and Carduus species by Freese (1994), C. litura was found only in Canada thistle. The stem- and petiole-galling fly Urophora cardui (L.) (Diptera: Tephritidae) is reported in the field in Europe only from Canada thistle (Zwölfer, 1965a) and the closely related species Cirsium setosum von Bieberstein (sometimes treated as a synonym of C. arvense) (Frenzel et al., 2000). It was screened by Peschken and Harris (1975) against 14 other European Cardueae species and against 11 species, mainly economically important plants, in other tribes and families. In these tests, consistent oviposition was seen only on Canada thistle, with occasional oviposition on Cirsium vulgare (Savi) Ten. and Carduus acanthoides L. Host specificity tests on the weevil L. planus were conducted by McClay (1989), who found that L. planus will not feed on ornamental or economic species in the tribe Cardueae and that Canada thistle is preferred over other Cirsium species for feeding and oviposition. These tests suggested that small-flowered Cirsium species were more suitable as hosts than native Cirsium species, which generally have larger flower heads. However, Louda and O‘Brien (2002) found L. planus reducing seed production of the large-flowered native Cirsium undulatum (Nutt.) Spreng. var. tracyi (Rydb.) Welsh in Colorado, indicating that redistribution of this insect poses greater risks to native species than was previously believed. Cassida rubiginosa is recorded from numerous species of Arctium, Carduus, Cirsium, Silybum, Onopordum, and Centaurea. In feeding tests, adults and larvae accept species from all these genera, as well as from globe artichoke (Zwölfer and Eichhorn, 1966; Zwölfer, 1969). Spring and Kok (1997) found that C. rubiginosa shows no oviposition preference between Canada thistle and Carduus thoermeri Weinmann; however, mortality of immature stages is lower on Canada thistle. They also observed adults, larvae, and egg masses on burdock, Arctium minus (Hill) Bernhardi, in the field, and reared C. rubiginosa from egg to adult on this species. Host specificity testing also was conducted on the lace bug Tingis ampliata Herrich-Schäffer (Hemiptera: Tingidae) (Peschken, 1977a) and the leaf beetle Lema cyanella (L.) (Coleoptera: Chrysomelidae) (Peschken and Johnson, 1979; Peschken, 1984b). Tingis ampliata was never released in North America because of concerns about possible attack on globe artichoke, Cynara scolymus L. Limited releases of L. cyanella have been made in Canada but no further releases or redistribution are planned (McClay, unpub. data). Cleonis pigra attacks numerous species of Cardueae in Europe, and is an economic pest of globe artichoke (LaFerla, 1939; Zwölfer, 1965a; Batra et al., 1981). Terellia ruficauda has been reared from six Cirsium species in Europe (Zwölfer, 1965a). Releases Made Information on releases of biological control agents against Canada thistle was obtained from the literature and, for the period between 1981 and 1985, from the USDA, ARS database on natural enemy releases in the United States (ROBO at http://www.ars-grin.gov/nigrp/robo.html). There undoubtedly have been many additional releases that have not been published; for example, 18 releases of C. litura and 12 of U. cardui were made in the eastern part of South Dakota between 1987 and 1984, and L. planus and C. rubiginosa also have been released in this area (R. Moehringer, S. Dakota Dept. of Agriculture, pers. comm.). Releases of A. carduorum began in 1966, using material from Switzerland via Canada, and were made in Delaware, Indiana, Maryland, Minnesota, New Jersey, South Dakota, and Wisconsin; in 1970, material from France was released in Maryland, New Jersey, and South Dakota (Julien and Griffiths, 1998). Two releases of A. carduorum from a population collected near Rome, Italy, were made in Maryland in 1982. Releases of C. litura began in 1971 (Julien and Griffiths, 1998). This weevil was released in Maryland on 16 occasions from 1982 to 1985 and at one site in New York State in 1984. Most of these releases were made using material imported from Bavaria, Germany, but four releases were made with material from established field populations in Montana. Urophora cardui was released on nine occasions in Maryland between 1981 and 1984, mostly using material from field collections near Vienna, Austria. Two releases of U. cardui from this source also were made in Iowa in 1982 and 1985. Another series of four releases totaling 4,400 adults from the population in British Columbia, Canada, was made in 1985 in Virginia (Kok, 1990). Cassida rubiginosa was moved from northern Virginia to a southwestern area of the state where it previously had not occurred (Ward and Pienkowski, 1978a). Biology and Ecology of Key Natural Enemies Altica carduorum Guérin-Méneville (Coleoptera: Chrysomelidae) This species has a Mediterranean and partly Atlantic distribution in Europe (Zwölfer, 1965b). A closely related species, Altica cirsicola Ohno, occurs in China and Japan (Laroche et al., 1996); however, RAPD fingerprinting (a DNA identification method) showed that a population from Xinjiang in western China was A. carduorum (Wan and Harris, 1995). Thus the distribution of A. carduorum extends from the Mediterranean and eastern Europe, through Kazakhstan, Kirghizia and Tadzhikistan to western China (Wan and Harris, 1995). In Switzerland, overwintering adults of A. carduorum begin to appear on foliage of Canada thistle in mid-April and oviposition starts in early May. Larvae are present on the leaves from mid-May through late July. Newly emerged adults feed heavily on foliage in August and September before leaving the plant to seek overwintering sites (Zwölfer, 1965b). Females oviposit on the underside of Canada thistle leaves, usually laying about 12 eggs per day. Under laboratory conditions, eggs hatch in about 11 days, larval development requires about one month, and pupa develop to adults in 10 to 11 days. Larvae feed on the undersurface of leaves, producing “windows” of clear epidermis. Adult feeding damage is evenly dispersed over the whole plant; heavy adult feeding can cause the collapse of plants both in the laboratory and in the field (Karny, 1963). Cassida rubiginosa Müller (Coleoptera: Chrysomelidae) This univoltine shield beetle feeds on foliage of several Cardueae species, both as adults and larvae. In Virginia, adults appear in late winter and oviposit, mainly on the underside of thistle leaves, from mid-March to early July. Eggs are laid in oothecae containing about five eggs. Development from egg to adult requires 435 degree-days above a threshold of 10.4°C. New generation adults begin to appear in late spring and can be found on plants up to November. Females produce an average of 815 eggs under laboratory conditions (Ward and Pienkowski, 1978a). In the field, C. rubiginosa is attacked by several larval parasitoids including Tetrastichus rhosaces (Walker) (Hymenoptera: Eulophidae) and Eucelatoriopsis dimmocki (Aldrich) (Diptera: Tachinidae) (Ward and Pienkowski, 1978b). However, Ang and Kok (1995) felt that parasitism did not limit C. rubiginosa populations in Virginia. Tipping (1993) found that adults released on Canada thistle in Maryland remained in close proximity to the release point and that most oothecae were laid within 1.6 m of the release point. Parasitism in this study was 10.5%, with the most common parasitoid being E. dimmocki. Larvae and pupae are heavily predated by larvae of Coccinella septempunctata L. (Coleoptera: Coccinellidae) in Maryland (P. Tipping, pers. comm.). Spring and Kok (1999) found about 21% overwintering survival of adult C. rubiginosa. Ceutorhynchus litura (F.) (Coleoptera: Curculionidae) This stem- and root-mining weevil occurs in France, Switzerland, Austria, Germany, Britain, and southern Scandinavia (Zwölfer and Harris, 1966) (Fig. 3). Females oviposit into the mid-veins of rosette leaves of Canada thistle leaves in spring. Eggs are laid in groups of one to five in a cavity made with the rostrum in the underside of a young leaf. Larvae hatch after five to nine days and mine down through the vein into the base of the stem and upper tap root (Fig. 4). There they form a feeding tunnel that may cause the stem to become somewhat inflated into an indistinct gall. Mature larvae leave the stem and pupate in a cocoon of soil particles, from which they emerge in late summer (Zwölfer and Harris, 1966; Peschken and Beecher, 1973). Adults overwinter in the soil or leaf litter.
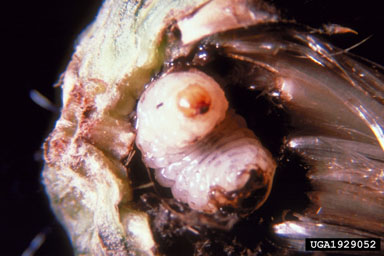
Cleonis pigra (Scopoli) (Coleoptera: Curculionidae) Adults of this large weevil emerge from overwintering sites in May and feed on Canada thistle foliage in June and July. The females oviposit into the lower portions of Canada thistle stems. The larvae mine down through the stem base into the root, which develops a spindle-shaped gall around the feeding site. Pupation occurs in the root, and adults emerge in late summer or fall (Anderson, 1956). Larinus planus (F.) (Coleoptera: Curculionidae) Adults of this weevil (Fig. 5) feed on Canada thistle foliage, but generally cause little damage. Females oviposit into the unopened flower buds, where larvae feed on the developing achenes and receptacle tissue. Larvae pupate in a cocoon formed of chewed host plant tissue (Fig. 6.). Only one larvae can complete development in each head. Adults emerge in late summer and overwinter in the litter (McClay, 1989).
Puccinia punctiformis Strauss (Röhling) (Uredinales: Pucciniaceae) This fungus is an autecious brachycyclic rust that produces systemic infections on the spring-emerging shoots of Canada thistle. Systemically infected shoots are pale and die before flowering, but spermogonia and uredosori are formed before plant death. Later in the season, infection of other shoots by uredospores leads to local infection followed by the formation of teliospores in autumn (Van Den Ende et al., 1987). Teliospores are responsible for the systemic form of infection (Van Den Ende et al., 1987; French and Lightfield, 1990). Germination of teliospores is stimulated by volatile compounds from germinating Canada thistle seeds and root cuttings (French et al., 1988; French et al., 1994). The germination rate of teliospores is highest in the temperature range of 10°C to 15°C (Frantzen, 1994). Systemic infection may be induced in the laboratory in root buds or seeds inoculated with teliospores (French and Lightfield, 1990; French et al., 1994), but it is not yet clear how root infection from teliospores could take place in the field (French et al., 1994). Systemically infected shoots are taller than uninfected ones but fail to flower and their root biomass is reduced (Thomas et al., 1994). Cumulative mortality of infected shoots in a field study in Maryland was 80% compared with less than 10% for healthy shoots (Tipping, 1993). Terellia ruficauda (F.) (Diptera: Tephritidae) Ovipositing females of this fly select female Canada thistle flower heads one day away from blooming. Eggs are laid between immature florets and the larvae feed on developing achenes through a hole drilled in the pericarp. Third instar larvae form cocoons of pappus hairs in which they overwinter; pupation and emergence take place in the spring (Lalonde and Roitberg, 1992). Urophora cardui (L.) (Diptera: Tephritidae) This univoltine stem-galling fly oviposits in the axillary buds of Canada thistle (Fig. 7). The eggs hatch in seven to 10 days. Larvae induce development of multi-chambered galls in the form of a swelling in the stem up to 23 mm in diameter (Lalonde and Shorthouse, 1985) (Fig. 8). Pupation and overwintering
Evaluation of Project Outcomes Establishment and Spread of Agents There is no evidence that A. carduorum has become established in the United States. Peschken (1977b) attributed the failure of this species to establish in Canada to predation. Ceutorhynchus litura is reported to be established in Maryland, North Dakota, South Dakota, and Virginia (Julien and Griffiths, 1998; P. Tipping, pers. comm.). Urophora cardui is reported to be established in Maryland and Virginia (Julien and Griffiths, 1998), although the species is probably not currently established in Maryland (P. Tipping, pers. comm.). Galls of U. cardui were found at all Virginia field sites visited in 1986 but at only two sites in 1987; however, numbers of galls had increased at these two sites in 1987 (Kok, 1990). No recoveries are recorded in Virginia since 1987. Urophora cardui is common along the Hudson River and in other areas in New York state (B. Blossey, pers. comm.). Its establishment status in Iowa is unknown. C. rubiginosa became established in southwestern Virginia and has persisted there for more than 20 years (Ang and Kok, 1995). Suppression of Target Weed There has been little work done to evaluate the effectiveness of biological control agents for Canada thistle in the eastern United States. Using Canada thistle root cuttings transplanted to caged field plots, Ang et al. (1995) showed that feeding by C. rubiginosa significantly reduced biomass and survival of Canada thistle. The effects of C. rubiginosa were stronger than those of plant competition from tall fescue (Festuca arundinacea Schreb.) and crownvetch (Coronilla varia L.). Similar results were obtained by Bacher and Schwab (2000) in Switzerland. Forsyth and Watson (1986) evaluated the stress inflicted on Canada thistle by four insect species and one pathogen in Québec, Canada. The seed head predator O. ruficauda reduced seed production by about 22%. Root mining by C. pigra sometimes killed plants. Main shoot galling by U. cardui reduced plant height and number of flowers, but side-shoot galling had less impact. Reports of the impact of C. litura have been varied. Based on field sampling, Rees (1990) suggested that this species had a significant impact on survival of Canada thistle in Montana. Peschken and Derby (1992), however, found in controlled experiments that combined attack by this species and U. cardui had no significant effect on most performance parameters of Canada thistle. The impact of biological control of Canada thistle in terms of economic benefits or recovery of native plant communities has not been evaluated. Recommendations for Future Work Future Needs for Importation or Evaluation The impact of currently established agents needs to be evaluated by controlled experimental methods, preferably using naturally-occurring densities of agents in field weed stands (McClay, 1995). The extent of non-target damage from the currently established agents also need to be further assessed. Such damage has been shown repeatedly for the seed weevil R. conicus, released as a biocontrol agent for Carduus and Silybum species (Rees, 1977; Louda, 1999; Herr, 2000). As with R. conicus, most of the agents released against Canada thistle have laboratory host ranges that include many native Cirsium species, but it is not known whether any of these native species are in fact being damaged, or are at risk of damage, in the field. Information on this would provide a valuable test of the reliability of laboratory host-range tests in predicting non-target utilization in the field. The European range of Canada thistle has been extensively explored for potential biocontrol agents, and it seems unlikely that there are promising agents yet undiscovered in this region. Further exploration in Central Asia and China may identify other possible candidate agents, and such exploration is planned (A. Gassmann, pers. comm.). Other Comments Canada thistle may be a difficult target for biological control for two reasons. Firstly, it is a significant agricultural weed in its native range in Europe (Schroeder et al., 1993), suggesting that its natural enemies there are not very effective in limiting its population density, at least under agricultural conditions. There has been little study of the impact of herbivory on natural populations of Canada thistle in Europe; however, Edwards et al. (2000) found that exclusion of insects with chemical pesticides had no effect on recruitment or density of Canada thistle in cultivated soil or grassland in southern England. Secondly, Canada thistle is congeneric with a large number of native North American Cirsium species, raising concerns about non-target damage to native species by introduced biological control agents (Louda, 1999; Louda and O‘Brien, 2002). Although some phytophagous insects associated with Canada thistle, such as U. cardui, appear to be virtually monophagous, others have a broad host range within the genus Cirsium and also will accept species of Carduus or related genera. In the past, several agents have been approved for release against Canada thistle on the basis of host specificity tests that would not be considered sufficient justification for release today. Future progress in classical biological control of Canada thistle will depend on the identification of new, adequately host specific herbivores from its native range, and will require improvements in host-testing procedures to allow better prediction and evaluation of non-target impacts. Acknowledgments I thank Philip Tipping, André Gassmann, Dean Kelch, Charles O‘Brien, and Ron Moehring for comments on the manuscript and for providing unpublished information. References Anderson, D. M. 1956. Notes on Cleonus piger (Scop.) in the United States (Coleoptera, Curculionidae). Anderson, R. S. 1987. Systematics, phylogeny and biogeography of New World weevils of the tribe Ang, B. N. and L. T. Kok. 1995. Seasonal mortality of Cassida rubiginosa (Coleoptera, Chrysomelidae) Ang, B. N., L. T. Kok, G.I. Holtzman, and D.D. Wolf. 1995. Canada thistle [Cirsium arvense (L.) Scop] Bacher, S. and F. Schwab. 2000. Effect of herbivore density, timing of attack and plant community on Batra, S. W. T., J. R. Coulson, P. H. Dunn, and P. E. Boldt. 1981. Insects and fungi associated with Berestetsky, A. O. 1997. Mycobiota of Cirsium arvense and allied species over the territory of the Bremer, K. 1994. Asteraceae: Cladistics and Classification. Timber Press, Portland, Oregon, USA. Donald, W. W. 1994. The biology of Canada thistle (Cirsium arvense). Reviews of Weed Science 6: 77- Donald, W. W. and M. Khan. 1996. Canada thistle (Cirsium arvense) effects on yield components of Edwards, G. R., G. W. Bourdôt, and M.J. Crawley. 2000. Influence of herbivory, competition and soil Forsyth, S. F. and A. K. Watson. 1986. Stress inflicted by organisms on Canada thistle, pp. 425-431. Frantzen, J. 1994. The effect of temperature on the germination of teliospores of Puccinia punctiformis. Freese, G. 1994. The insect complexes associated with the stems of seven thistle species. French, R. C. and A. R. Lightfield. 1990. Induction of systemic aecial infection in Canada thistle French, R. C., S. K. Turner, P. E. Sonnett, P. Pfeffer, and E. Piotrowski. 1988. Properties of an extract French, R. C., S. E. Nester, and R. G. Binder. 1994. Volatiles from germinating Canada thistle seed Frenzel, M., S. Eber, S. Klotz, and R. Brandl. 2000. Ecological comparisons across geographical Harris, P. 1964. Host specificity of Altica carduorum Guer. (Coleoptera: Chrysomelidae). Canadian Herr, J. C. 2000. Evaluating non-target effects: the thistle story, pp. 12-17. In Hoddle, M. (ed.). Hiebert, R. D. and J. Stubbendieck. 1993. Handbook for Ranking Exotic Plants for Management and Hunter, J. H. and L. W. Smith. 1972. Environment and herbicide effects on Canada thistle ecotypes Jessep, C. T. 1989. Cirsium arvense (L.) Scopoli, Californian thistle (Asteraceae), pp. 343-345. In Jewett, J. G., C. C. Scheaffer, R. D. Moon, N. P. Martin, D. K. Barnes, D. D. Breitbach, and N. R. Johnson, D. R., D. L. Wyse, and K. J. Jones. 1996. Controlling weeds with phytopathogenic bacteria. Julien, M. H. and M. W. Griffiths (eds.). 1998. Biological Control of Weeds: A World Catalogue of Karny, M. 1963. Haltica carduorum Guer. (Col.: Chrysomelidae) attacking Cirsium arvense. Technical Kok, L. T. 1990. Biological control of weeds in Virginia from 1969-86, pp. 623-629. In Delfosse, E. S. LaFerla, A. 1939. Contributo alla conoscenza dei cleono del carciofo (Cleonus piger Scop.). Bolletino Lalonde, R. G. and B. D. Roitberg. 1992. Field studies of seed predation in an introduced weedy thistle. Lalonde, R. G. and J. D. Shorthouse. 1985. Growth and development of larvae and galls of Urophora Laroche, A., R. A. De Clerck-Floate, L. Lesage, K. D. Floate, and T. Demeke. 1996. Are Altica Lloyd, D. G. and A. J. Myall. 1976. Sexual dimorphism in Cirsium arvense (L.) Scop. Annals of Botany Louda, S. M. 1999. Negative ecological effects of the musk thistle biological control agent, Rhinocyllus Louda, S.M. and C.W. O’Brien. 2002. Unexpected ecological effects of distributing the exotic weevil, McClay, A. S. 1989. The potential of Larinus planus (F.) (Coleoptera: Curculionidae), an accidentally McClay, A. S. 1995. Beyond “before-and-after”: experimental design and evaluation in classical weed McClay, A. S., R. Bourchier, R. A. Butts, and D. P. Peschken. 2001. Cirsium arvense (L.) Scopoli, McLennan, B. R., R. Ashford, and M. D. Devine. 1991. Cirsium arvense (L.) Scop. competition with Moore, R. J. 1975. The biology of Canadian weeds. 13. Cirsium arvense (L.) Scop. Canadian Journal of Moyer, J. R., G. B. Schaalje, and P. Bergen. 1991. Alfalfa (Medicago sativa) seed yield loss due to Nadeau, L. B. and W. H. Vanden Born. 1989. The root system of Canada thistle. Canadian Journal of Nuzzo, V. 1997. Element Stewardship Abstract for Cirsium arvense. The Nature Conservancy, O‘Brien, C. W. and G. J. Wibmer. 1982. Annotated checklist of the weevils (Curculionidae sensu lato) O’Sullivan, P. A., V. C. Kossatz, G. M. Weiss, and D. A. Dew. 1982. An approach to estimating yield O’Sullivan, P. A., G. M. Weiss, and V. C. Kossatz. 1985. Indices of competition for estimating Peschken, D. P. 1977a. Host specificity of Tingis ampliata (Tingidae: Heteroptera): a candidate for the Peschken, D. P. 1977b. Biological control of creeping thistle (Cirsium arvense): analysis of the releases Peschken, D. P. 1984a. Cirsium arvense (L.) Scop., Canada thistle (Compositae), pp. 139-146. In Peschken, D. P. 1984b. Host range of Lema cyanella (Coleoptera: Chrysomelidae), a candidate for Peschken, D. P. and R. W. Beecher. 1973. Ceutorhynchus litura (Coleoptera: Curculionidae): biology Peschken, D. P. and J. L. Derby. 1992. Effect of Urophora cardui (L.) (Diptera: Tephritidae) and Peschken, D. P. and P. Harris. 1975. Host specificity and biology of Urophora cardui (Diptera: Peschken, D. P. and G. R. Johnson. 1979. Host specificity and suitability of Lema cyanella Redfern, M. 1983. Insects and Thistles. Cambridge University Press, New York. Rees, N. E. 1977. Impact of Rhinocyllus conicus on thistles in southwestern Montana. Environmental Rees, N. E. 1990. Establishment, dispersal and influence of Ceutorhynchus litura on Canada thistle ( Schroeder, D. 1980. The biological control of thistles. Biocontrol News and Information 1: 9-26. Schroeder, D., H. Müller-Schärer, and C. S. A. Stinson. 1993. A European weed survey in 10 major Skinner, K., L. Smith, and P. Rice. 2000. Using noxious weed lists to prioritize targets for developing Spring, A. and L. T. Kok. 1997. Field studies on the reproductive capacity and development of Cassida Spring, A. and L. T. Kok. 1999. Winter survival of Cassida rubiginosa (Coleoptera: Chrysomelidae), a Stachon, W. J. and R. L. Zimdahl. 1980. Allelopathic activity of Canada thistle (Cirsium arvense) in Stary, P. 1986. Creeping thistle, Cirsium arvense, as a reservoir of aphid parasitoids (Hymenoptera, Thomas, R. F., T. J. Tworkoski, R.C. French, and G.R. Leather. 1994. Puccinia punctiformis affects Tipping, P.W. 1992. Density of Carduus and Cirsium thistles in selected areas of Maryland. Weed Tipping, P.W. 1993. Field studies with Cassida rubiginosa (Coleoptera: Chrysomelidae) in Canada USDA, NRCS (United States Department of Agriculture, Natural Resources Conservation Service). Van Den Ende, G., J. Frantzen, and T. Timmers. 1987. Teleutospores as origin of systemic infection of Volkl, W. 1989. Resource partitioning in a guild of aphid species associated with creeping thistle Wan, F.-H. and P. Harris. 1995. Biology, host specificity and suitability of the Wusu, NW China biotype Wan, F. H. and P. Harris. 1996. Host finding and recognition by Altica carduorum, a defoliator of Wan, F. H. and P. Harris. 1997. Use of risk analysis for screening weed biocontrol agents – Altica Wan, F.H., P. Harris, L.M. Cai, and M.X. Zhang. 1996. Host specificity of Altica carduorum Guer. Ward, R. H. and R. L. Pienkowski. 1978a. Biology of Cassida rubiginosa, a thistle-feeding shield beetle. Ward, R. H. and R. L. Pienkowski. 1978b. Mortality and parasitism of Cassida rubiginosa, a thistle- Wheeler, A. G., Jr. and D. R. Whitehead. 1985. Larinus planus (F.) in North America (Coleoptera: Winiarska, W. 1986. Roslinozerna entomofauna ostrozenia polnego, Cirsium arvense (L.) Scop. Youssef, N. N. and E. W. Evans. 1994. Exploitation of Canada thistle by the weevil Rhinocyllus conicus Zwölfer, H. 1965a. Preliminary list of phytophagous insects attacking wild Cynareae (Compositae) in Zwölfer, H. 1965b. Observations on the distribution and ecology of Altica carduorum Guer. (Col. Zwölfer, H. 1969. Experimental feeding ranges of species of Chrysomelidae (Col.) associated with Zwölfer, H. and O. Eichhorn. 1966. The host ranges of Cassida (Col.: Chrysomelidae) species attacking Zwölfer, H. and P. Harris. 1966. Ceutorhynchus litura (F.) (Col., Curculionidae), a potential insect for the [ Contents ] [ Previous ] [ Next ] |
|
| ||||
| |
The Bugwood Network and Forestry Images Image Archive and Database Systems The University of Georgia - Warnell School of Forestry and Natural Resources and College of Agricultural and Environmental Sciences - Dept. of Entomology Last updated on Thursday, July 19, 2018 at 01:38 PM Questions and/or comments to the | |||

![Figure 1. Canada thistle [Cirsium arvense (L.) Scopoli]. (Photograph by L.M. Dietz.)](https://bugwoodcloud.org/images/384x256/1929057.jpg)
![Figure 2. Canada thistle [Cirsium arvense (L.) Scopoli] infestation in canola (Brassica rapa L.). (Photograph by A.S. McClay.)](https://bugwoodcloud.org/images/384x256/1929056.jpg)